

Abstract
B cells play a crucial role in immunity against various infectious diseases. However, their role in tuberculosis (TB) has been largely understudied. Emerging evidence suggests that B cells actively shape immune responses in TB. Beyond their classical functions, B cells contribute to the formation of inducible bronchus-associated lymphoid tissue (iBALT), a tertiary lymphoid structure (TLS) that enhances localized immune responses in the lungs. As iBALT is a site for B-T cell interactions and the generation of high-affinity antibodies, recent studies suggest that sex differences in iBALT formation influence TB immunity. This review synthesizes evidence from both TB and non-TB models to highlight the expanding role of B cells and iBALT, underscoring their potential implications for vaccine development and immunotherapy.
1 Introduction
Tuberculosis (TB), caused by Mycobacterium tuberculosis (Mtb), remains a leading cause of infectious disease-related mortality worldwide (1). Despite extensive research efforts, developing an effective vaccine beyond Bacille Calmette-Guérin (BCG) continues to face substantial hurdles (2, 3). This challenge is partly due to the limited understanding of the mechanisms underlying protective immunity and the distinction between beneficial and pathogenic immune responses (4, 5). Given the intracellular nature of Mtb, TB research has traditionally centered on T cell-mediated immune responses (6–10). While T cells are undoubtedly essential, emerging evidence highlights the previously underappreciated role of B cells in TB immunity (11–16). Beyond their classical function of antibody production, B cells contribute significantly to antigen presentation (17), cytokine secretion (18, 19), and the formation of tertiary lymphoid structures (TLS), such as inducible bronchus-associated lymphoid tissue (iBALT) (16, 20–23). Notably, recent studies also suggest that the formation and function of iBALT can exhibit sex-specific differences, potentially influencing the progression and outcome of TB disease (24–27). Intriguingly, males are more susceptible than females according to global statistics. Sex hormones along with genetic factors, drive differences in immune responses between males and females, which may shape iBALT organization and its protective efficacy against Mtb infection (27). These findings suggest that B cells are not mere bystanders but central players in TB pathogenesis and protection, necessitating a reassessment of their role in shaping immune responses to Mtb infection, particularly in both sexes.
Historically, evaluating immune responses in TB, particularly the diversity and functional heterogeneity of B cells, their interactions with other immune cells, and their specific roles within the lungs—the primary site of infection—has been challenging. However, recent advancements in immunological techniques, including spatial transcriptomics and single-cell RNA sequencing, now allow for a more detailed characterization of the tissue microenvironment and the identification of distinct subsets of lung-resident B cells (14, 28, 29). This represents a new opportunity to unravel the role of B cells in localized immune response to Mtb infection; knowledge that will improve our understanding of the immune correlates of protection in TB and may inform the development of innovative therapeutic strategies. In this review we summarize these emerging insights, address current knowledge gaps and encourage deeper exploration into the diverse functions of B cells in TB immunity.
2 B cells and TB: what we know so far
Traditionally, B cells have been regarded as secondary players in the immune response to Mtb. However, over the past decade, numerous studies have challenged this view, revealing a potentially pivotal role for B cells in TB immunity (23, 30–33). During Mtb infection, B cells are reduced in peripheral blood and present within the granulomas in the lung (14, 32, 34–37), the hallmark structures of TB pathology. Moreover, B cells are well recognized for their canonical roles in immunity, such as antibody production (38); however, studies over the past decade have uncovered non-canonical functions of B cells that further expand their immunological significance (39). Through antigen presentation, cytokine production, and antibody secretion, B cells shape the local immune microenvironment and influence the dynamics of host-pathogen interactions (40, 41).
A particularly intriguing aspect of B cell activity in TB is their presence in TLS/iBALT. These structures emerge in response to chronic inflammation or infection and provide a localized lung environment where adaptive immunity is enhanced (42, 43). Here B cells also function as antigen-presenting cells (APCs), interacting with T cells to amplify immune responses (40, 44, 45). This underscores their importance in bridging humoral and cellular immunity to effectively combat Mtb.
A comprehensive understanding of both canonical and non-canonical roles is crucial to fully unravel the complexity of B cell-mediated immunity in TB. Here, we consolidate current knowledge on the diverse functions of B cells in TB and other infectious and non-infectious diseases, highlighting both established and emerging roles, with a focus on iBALT.
2.1 Canonical role of B cells
Antibodies can directly neutralize Mtb by binding to its surface antigens, thereby preventing bacterial adhesion to host cells and intracellular entry (30). Through opsonization, antibody-coated Mtb is internalized by macrophages via Fc receptor-mediated uptake, known as antibody-dependent cellular phagocytosis (ADCP), a process that promotes phagosome maturation and enhances bacterial clearance (46, 47). Studies have identified anti-lipoarabinomannan (LAM) antibodies as particularly effective in inhibiting Mtb growth within macrophages by enhancing opsonization and promoting phagosome maturation (48, 49). Moreover, these antibodies have been shown to promote bacterial clearance through complement activation (50, 51). Fc regions also induce antibody-dependent cellular cytotoxicity (ADCC) reactions through neutrophil and natural killer (NK) cell degranulation which eliminates infected cells and controls bacterial load (47). Additionally, antibodies against Mtb-secreted proteins, including Ag85, ESAT-6, and CFP-10, are thought to disrupt bacterial virulence mechanisms and enhance host defense. Intriguingly, individuals highly exposed to Mtb who did not develop classical latent TB infection (as defined by positive IFN-γ release assay or tuberculin skin test) exhibited Mtb-specific humoral immunity with enhanced avidity, a shift toward the IgG1 subclass, and distinct IgG Fc-glycosylation profiles, instead of the typical IFN-γ response (52). Importantly, differential Fc glycosylation was found to modulate Fc-mediated antibody activity, indicating its potential to directly impact Mtb control (30). Furthermore, the corresponding Fc receptors (FcRs) appear to shape the overall immune response to Mtb. Mice lacking activating FcRs develop more severe immunopathology and higher bacterial burdens, whereas mice deficient in the inhibitory FcR show reduced pathology (53). Collectively, these lines of evidence suggest that Fc-mediated antibody effector functions together with the expression of Fc receptors significantly contribute to the modulation of TB disease.
Despite these advances, the precise contribution of antibodies to protection against TB remains contentious. Passive transfer experiments have shown that antibodies can confer measurable protection in some settings (31, 54–56), yet other studies report minimal or no benefit, highlighting the complexity of humoral responses to Mtb (33, 57). The protective capacity of antibodies likely depends on multiple factors, including isotype, antigen specificity, and Fc receptor engagement. Recent developments in systems serology and high-throughput sequencing now enable detailed profiling of the TB-specific antibody repertoire, offering new insight into how distinct antibody subclasses and functions may contribute to protection.
2.2 Non-canonical role of B cells
While B cells are best known for their role in antibody-mediated immunity, they also perform several non-canonical, regulatory, or tissue-specific functions, which are increasingly recognized. Among these roles, cytokine production has emerged as a critical mechanism by which B cells shape both innate and adaptive responses. A specialized subset known as regulatory B cells (Bregs) is particularly enriched for IL-10 secretion, enabling these cells to suppress excessive inflammation and maintain immune homeostasis (58, 59). Bregs have been detected in the blood of individuals with active TB (58), and they are also present in the lungs of Mtb-infected mice, particularly during the chronic phase of infection (59). Recent findings indicate that the absence of B cell-derived IL-10 enhances resistance to TB, suggesting that B cell–specific IL-10 interferes with optimal infection control (24). B cells can also produce other immunoregulatory cytokines that may impact TB outcomes including IL-35, and IL-35/IL-20 co-producing B cells have been identified in TB (60). Complementing these observations, in vivo studies have shown that B cell derived IL-35 can suppress T cell responses, potentially undermining anti-Mtb immunity (61). Likewise, type I IFN producing B cells may contribute to impaired Mtb control by promoting macrophage polarization towards an anti-inflammatory phenotype (62). Conversely, Tbet expressing B cell can secrete IFN-γ, which is central to TB immunity, and marginal zone B cells secrete numerous cytokines in response to Mtb stimulation ex vivo, including TNF-a, IL-2 and GM-CSF (63). Although these cytokines play an important role in TB the relevance of their B cell source in vivo is not yet clear. Finally, Linge et al. also demonstrate that B cell-derived IL-6 production following Mtb infection exerts pleiotropic effects on both B and T cell function (64). Collectively, these findings indicate that cytokine-producing B cell subsets can exert both protective and potentially detrimental effects during Mtb infection, shaping the balance between host defense and pathogen persistence.
One of the most intriguing aspect of B cells in TB is their participation in the formation of iBALT, a type of TLS that develops in the lung (40). Unlike secondary lymphoid organs (SLOs) such as lymph nodes, spleen, tonsils, and Peyer’s patches, which are anatomically predetermined, TLS develop dynamically in response to localized immune challenges (65–67). These structures form directly at the site of Mtb infection, creating a highly specialized immune microenvironment that promotes antigen presentation, cytokine signaling, and the maintenance of immune memory (40).
3 Putative mechanisms underlying iBALT formation: role of inflammation, infection, and cytokines
Although the precise mechanisms underlying iBALT formation in TB remain undefined, data from other respiratory infections indicate that inflammation, antigenic stimulation, lymphotoxin (LT) signaling, cytokines, and chemokines are key drivers of its development (68–71). Pro-inflammatory cytokines such as interleukin-17 (IL-17), interleukin-6 (IL-6), and interleukin-22 (IL-22) promote immune-cell recruitment, stromal cell activation, and the clustering of B and T cells into organized aggregates (72, 73). In parallel, LTα/β-LTβR interactions between lymphoid tissue inducer-like cells (including B cells) and stromal cells help imprint a lymphoid organizer phenotype on local fibroblasts and endothelial cells. Chemokines including CXCL13, CXCL12, CCL21, and CCL19 then guide lymphocyte migration and compartmentalization, fostering the emergence of follicular-like zones within iBALT that resemble germinal centers (GCs) in SLOs and are predicted to support antigen-driven selection, affinity maturation, and memory B cell formation (70, 74, 75).
Importantly, these LT cytokine-chemokine networks represent only one part of a broader, heterogeneous landscape of iBALT-inducing mechanisms. Studies in influenza, Pneumocystis infection, and LPS-driven neonatal inflammation show that iBALT can arise through both lymphotoxin-dependent and lymphotoxin-independent routes, with different cell types (e.g., Th17 cells, γδ T cells, ILC3, dendritic cells, and activated B cells) acting as lymphoid tissue inducer (LTi)-like populations, and with additional contributions from IL-1 family cytokines, type I interferons, and TNF-family members beyond LTα/β (76, 77). Moreover, iBALT may form de novo or via remodeling of pre-existing peribronchial/perivascular leukocyte-stromal aggregates, and the relative contribution of these pathways is likely shaped by pathogen type, antigen load, age, and the chronicity of inflammation (74, 76). Thus, the model below should be viewed as one experimentally supported framework rather than a single, exclusive mechanism of iBALT formation (Figure 1).
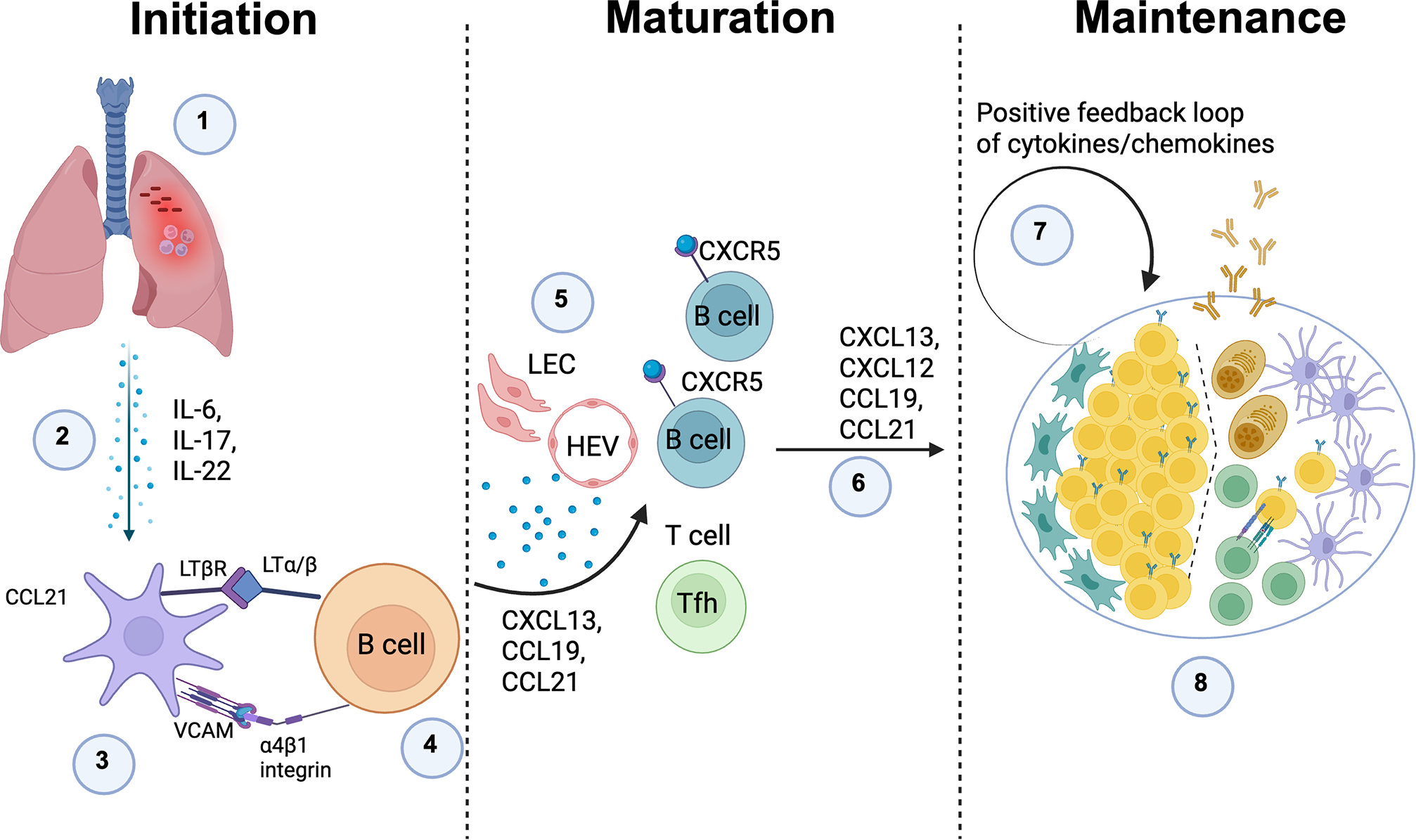
Putative mechanism of iBALT formation in TB: 1) Mtb infection leads to chronic inflammation; 2) Persistent inflammation drives the release of pro-inflammatory cytokines, including TNF family members, IL-6, IL-17, and IL-22, which promote immune cell recruitment and tissue remodeling; 3) Inflammatory cytokines activate lung stromal cells, prompting them to function as lymphoid tissue organizer (LTo) cells by upregulating LTβR and VCAM expression on their surface. This transformation is critical for TLS formation; 4) Activated B cells express LTα/β, which binds to LTβR on stromal cells, initiating their function as lymphoid tissue inducer (LTi) cells and promoting lymphoid-like structures; 5) LTα/β-LTβR interactions drive the expression of CXCL13, CCL19, and CCL21, which recruit B cells, T cells, and dendritic cells (DCs) to the inflamed site, creating a localized immune niche; 6) Production of CCL19, CCL21, CXCL12, and CXCL13, ensures the persistence of inducible bronchus-associated lymphoid tissue (iBALT) and reinforces local immune response; 7) A positive feedback loop amplifies the secretion of these critical chemokines and cytokines, stabilizing the iBALT structure and supporting its function as a persistent immune hub; 8) iBALT structures may develop germinal center-like formations, where T and B cells interact, leading to the generation of high-affinity antibodies against Mtb antigens, enhancing immune defense against infection. The image was created using Biorender. Define in the legend: Lymphatic endothelial cells (LEC), High endothelial venules (HEV).
3.1 Initiation
A local inflammatory insult in the lung for example infection induces production of cytokines such as IL-6, IL-17 and IL-22. These signals activate perivascular/peribronchial stromal cells, which begin to express CCL21, VCAM-1 and LTβR. Circulating B cells expressing α4β1 integrin and membrane LTα/β are recruited to these sites and, through VCAM-integrin and LTα/β-LTβR interactions, start to imprint a lymphoid-organizer phenotype on the stromal cells, seeding early TLS aggregates.
3.2 Maturation
As the niche stabilizes, lymphatic endothelial cells (LEC) and high endothelial venules (HEV) are induced, enabling efficient entry and exit of lymphocytes. Activated stromal cells, follicular dendritic cell (FDC) precursors and fibroblasts secrete homeostatic chemokines CXCL13, CXCL12, CCL19 and CCL21, which attract CXCR5+ and CXCR4+ B cells and CCR7+ T cells, including T follicular helper cells (Tfh). This chemokine-guided influx and segregation of cells drives the emergence of distinct B cell follicles and T cell zones, giving the structure a secondary-lymphoid-organ like architecture.
3.3 Maintenance
In the mature TLS, ongoing antigen stimulation and cytokine/chemokine production establish a positive feedback loop. GC like B cell clusters, plasma cells, FDC networks and T cells are maintained within the structure, allowing sustained local antibody production and T cell help. Depending on the inflammatory context, similar stromal and cytokine circuits can also stabilize alternative TLS configurations, so this model represents one major pathway rather than the only mechanism by which TLS can form and persist.
4 Pathogenic versus protective iBALT
Depending on the context iBALT can be either protective or contribute to pathology. Perhaps the simplest example is in organ transplantation, where immune regulation by iBALT confers transplant success (78–80) but pro-inflammatory iBALT results in rejection (78). Similarly, iBALT is associated with protection in conditions where inflammation is required such as in response to infection (74), or it can exacerbate inflammation in response to self-antigens contributing to autoimmunity.
4.1 Protective iBALT
In most infectious contexts, iBALT is associated with protection (15, 20, 21, 25, 81–88). This is partly due to the production of high-affinity antibodies within iBALT, which neutralize pathogens and limit their spread. These responses are enhanced upon secondary exposure in the presence of iBALT (89, 90). In mice infected with influenza, iBALT was protective even in mice completely lacking secondary lymphoid organs (91). Conversely, NHP infants do not form iBALT, resulting in a lower antibody response and increased pulmonary damage (92). Additionally, iBALT has been associated with reduced inflammation in viral (82) and bacterial infections (83, 84).
Modified Vaccinia Ankara (MVA) and Pneumocystis infections highlight the significance of iBALT as a priming site against unrelated pathogens (85–87). In influenza models, local memory exhibits broader specificity compared to lymph node-selected memory (88). Additionally, iBALT has been linked to positive cancer outcomes (93–95). TLS have been shown to enhance CD4+ and CD8+ T cell responses in cancer patients (93), and the role of B cell-mediated priming of protective T cell responses is an active area of investigation in cancer immunotherapy. This parallels the protective responses observed in TB models, where iBALT formation is associated with antigen-specific CD4+ and CD8+ T cell responses (96) (Figure 2).
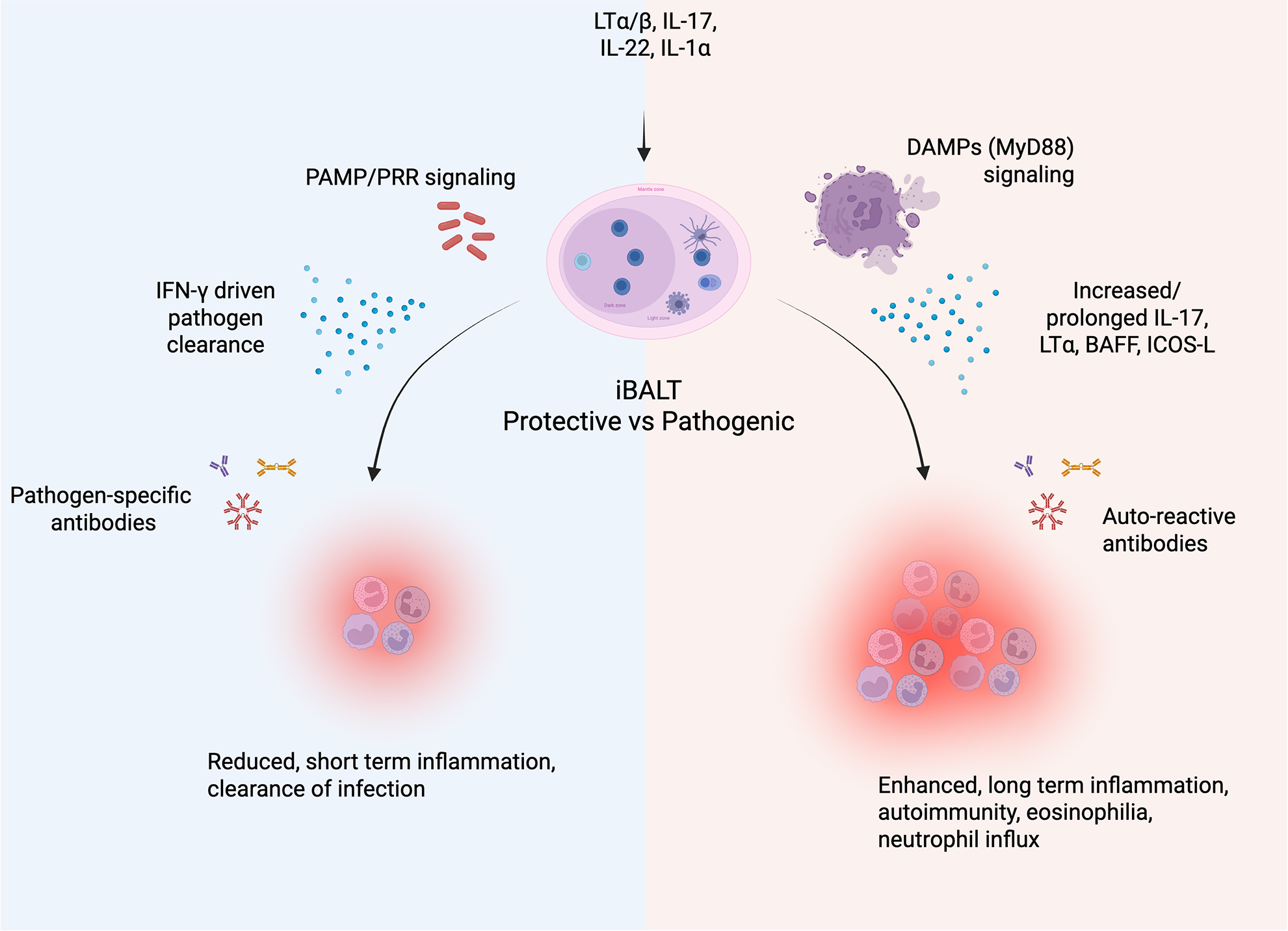
Description of protective vs pathogenic iBALT. Following the induction of iBALT, the local microenvironment can sway the response to be protective or pathogenic. In general, a protective response (blue) entails a specific response driven by PAMP/PRR signaling. This induces IFN-γ driven responses to clear the pathogen along with pathogen-specific antibodies, resulting in short-term inflammation. In contrast, a pathogenic response (red) is generally driven by DAMPs from local tissue damage accompanied by increased/prolonged IL-17, LTα, BAFF, ICOS-L, and auto-reactive antibodies. This leads to prolonged inflammation, eosinophilia, neutrophil influx, and autoimmune pathology. The image was created using Biorender.
4.2 Pathogenic iBALT
Induced hypersensitivity and persistent inflammation are often associated with detrimental iBALT formation. Continuous exposure to foreign substances, such as cigarette smoke, can induce lung inflammation, leading to chronic obstructive pulmonary disease (COPD). In this scenario, iBALT contributes to harmful inflammation associated with aging and inflammaging (97). Similarly, allergies involve persistent antigen stimulation, which can result in iBALT-associated immunopathology (98). Ongoing inflammation can also lead to a loss of tolerance and the development of autoimmune conditions such as Rheumatoid lung disease, Sjögren syndrome and pulmonary arterial hypertension (99). In these cases the sustained production of IL-17 and LT drives the formation of pathogenic iBALT (100). Consequently, elevated levels of BAFF, ICOS-L, and LTα drive B cell survival and auto-antibody production (75, 99, 101, 102), perpetuating the pro-inflammatory loop (Figure 2).
Respiratory syncytial virus (RSV) infection is a Th2 driven condition (103) where iBALT is linked to the disease (104). Interestingly, in Th2 and/or Th17 driven conditions (98, 105), allergies can further exacerbate RSV pathology (106). In graft versus host disease (GVHD), the recruitment of CXCR5+ Tfh and GC B cell worsens the disease (78), whereas recruitment of CXCR5+ Tregs reduces lung rejection (79). Notably, the recruitment of FoxP3+ Tregs into iBALT can prevent transplant rejection by inhibiting CD40 and CD28 signaling, and even permits multiple transplantations (80).
4.3 B cell heterogeneity as a driver of protective and pathogenic outcomes
Across infections, whether B cells are protective or pathogenic depends on which B cell subsets are engaged and the affinity/quality of the antibodies they produce. In well-organized GCs in lymph nodes, spleen, or iBALT, follicular and GC B cells undergo somatic hypermutation and stringent selection; high-affinity clones are positively selected into the memory and plasma-cell compartments, where they produce class-switched antibodies that efficiently neutralize pathogens and provide durable protection after infection or vaccination (107, 108). By contrast, extrafollicular plasmablasts and “atypical” or age-associated B cells often expand under conditions of high or persistent antigen load and strong inflammatory signals. In severe COVID-19, for example, dominant extrafollicular B cell responses are associated with robust but relatively low-mutation antibody production, lupus-like autoreactivity, and poor clinical outcomes, indicating how impaired affinity maturation can track with immunopathology (109). Bregs add another layer of complexity as IL-10/IL-35 producing Bregs can limit tissue damage in autoimmunity and some infections, but in chronic viral disease they may also suppress protective T cell responses and favor pathogen persistence (110, 111). In addition, as mentioned, marginal zone B cell, which are present in TB infected lung tissue (14), secrete many relevant cytokines following Mtb stimulation. Antibody quality then modulates these subset effects: high-affinity, strongly neutralizing antibodies from well-selected B cells are typically protective, whereas non- or sub-neutralizing antibodies generated by sub optimally selected B cells can form immune complexes or mediate antibody-dependent enhancement (ADE), as shown for dengue and other flaviviruses where intermediate titers of cross-reactive IgG can increase viral entry and drive severe disease via Fcγ-receptor–bearing myeloid cells (112).
Although detailed mechanistic data in TB are more limited, human and experimental studies already suggest a similar spectrum. B cell rich TLS/iBALT adjacent to granulomas contain GC-like B cells and Tfh cells and are often associated with better-organized lesions and evidence of local immune control, consistent with a protective role for well-selected, tissue-resident B cell responses and their antibodies (20, 113–115). Conversely, B cells can also promote granulomatous inflammation and lung damage in chronic murine TB, highlighting that dysregulated or chronically stimulated B cell compartments may tilt toward pathology (16). This protective versus pathologic balance likely depends not only on the presence of B cells, but also on how their activation is focused and selected within these local niches. This raises the possibility that Mtb actively perturbs B cell selection upstream of antigen specificity to limit adaptive immunity and/or drive inflammation and tissue destruction to enhance transmission. Several studies have shown that active TB leads to the expansion of antibodies targeting unrelated pathogens, including measles, RSV and tetanus (116, 117). The mechanism is unclear, but one intriguing possibility is the existence of superantigen-like immune modulation (118) by a TB derived product. This has been described for other bacterial pathogens and involves bacterial factors binding conserved immunoglobulin regions outside the complementarity determining regions (CDRs) to broadly engage BCRs and drive non-cognate activation, deletion, and repertoire distortion (119). Such broad BCR pressure could reshape the pool of B cells available for iBALT GC reactions and reduce the probability of generating high-affinity protective clones where they are needed most. However, while T cell-directed superantigen activity has been proposed in TB (118), no Mtb-derived B cell superantigen has been identified to date, and similar patterns of non-cognate B cell activation could also arise from alternative mechanisms such as innate receptor signaling and inflammatory cytokine milieus (120).
Taken together, these examples support a model in which (i) subset composition (GC vs extrafollicular vs regulatory vs tissue-resident B cells) and (ii) antibody affinity/function (neutralizing vs sub-neutralizing, pro-inflammatory Fc profiles) jointly determine whether B cell responses are net protective or pathogenic.
5 iBALT in TB
Accumulating evidence suggests a positive correlation between iBALT formation and improved control of Mtb and latent TB infection (LTBI) (81, 121, 122). The structural organization of iBALT facilitates close interactions between B cells, T cells, and APCs thereby promoting efficient antigen presentation and T cell priming. Specifically, B cells within iBALT can present Mtb antigens to CD4+ T cells, supporting their activation and differentiation into effector T cells that secrete IFN-γ and other cytokines essential for macrophage activation and bacterial clearance (40). This interaction between B cells and T cells within iBALT may significantly enhance immune efficacy, particularly in the early stages of Mtb infection when localized immune activation is critical for controlling bacterial replication (15, 40).
The formation of lymphoid follicles is observed not only in Mtb infection but also following mucosal vaccination, which confers superior protection against TB in animal models (22, 123–125). Non-human primate studies have shown that macaques vaccinated with MtbΔsigH demonstrate protective immunity, characterized by the absence of granulomas and the formation of iBALT, along with strong antigen-specific CD4+ and CD8+ T cell responses (22). Intriguingly, exposure to commensal gut bacteria confers protection to pneumonia in mice by priming ILC3 cells to migrate to the lung (126). ILC3s mediate an early protective role against Mtb infection by priming iBALT formation via IL-22 and IL-17 and thereby starting the CXCL13/CXCR5 cascade (127). Finally, oral exposure to non-tuberculous mycobacteria promotes iBALT formation in BCG vaccinated mice and subsequent protection (15). Similarly, delivery of Mtb antigen-primed dendritic cells (DCs) rapidly induced iBALT formation and near-sterilizing immunity in mice (128). Collectively, these findings highlight that the induction of local lymphoid structures and B cell responses at mucosal sites is a critical component of protective immunity against TB and represents a promising strategy for next-generation vaccines.
In humans, iBALT is linked to latency and containment of infection, whereas its absence is associated with active disease (81, 121). In NHP, the presence of iBALT is associated with protection (22, 129). Recently intravenous-BCG administration achieved complete protection in NHP, characterized by the distinct appearance of CD11c+ and CD3+ immune infiltrates, termed “microgranulomas”, which are associated with protection (96). A Th1/Th17 response also correlated with protection (130). An elevated TB-specific IgM response was observed in infected human lungs (14), and a robust IgM response was linked to protection in NHP (131). While B cell depletion had a modest effect on protection in this model, protection primarily relies on CD4 and CD8 immunity (132), again suggesting that iBALT may be an important mediator of protection.
While iBALT is generally associated with protective immunity in TB, it may also contribute to disease progression in certain cases (as described above), particularly when linked to chronic inflammation. A study by Chen et al. found B cells to enhance inflammation and host-detrimental immunopathology (16) by promoting Th1 responses while restricting anti-inflammatory IL-10. In the absence of B cells, mice survived Mtb infection significantly longer. Differences in study outcomes may be due, in part, to the use of different Mtb strains, as variations in pathogen characteristics—such as cell wall components and lipid moieties—can shape the host immune response and influence the balance between protective and harmful inflammation. Moreover, the formation and function of iBALT may be influenced by host factors such as genetic susceptibility, sex, age, or co-morbidities, all of which can impact the efficacy of the immune response. In particular, sex has been shown to have a profound impact on the immune system and may therefore significantly shape iBALT formation.
6 Sex differences in immunity and iBALT formation
Sex differences in the immune system arise from variations in the development, function, and regulation of diverse immune cell populations, shaping distinct responses in males and females. These differences are influenced by a combination of genetic, hormonal, and environmental factors and have implications for immunity, autoimmunity, and vaccine responses (133). The underlying reasons for sex differences in TB are not fully understood and were long thought to result from gender-related differences in exposure, behavior, and lifestyle (26, 27). The study of the role of biological sex in TB is relatively recent and is based, in part, on the observation that the male bias in TB incidence becomes apparent only after puberty (WHO report) —indirectly implicating sex hormones as potential key mediators of this sex difference. Indeed, growing experimental evidence supports a direct impact of biological sex on anti-TB immune responses. In particular, murine infection models, which allow the isolation of biological sex from gender-related confounders, consistently demonstrate increased susceptibility of males compared with females. Male mice exhibit significantly reduced survival following Mtb infection, and this phenotype is observed across different genetic backgrounds (e.g. C57BL/6 and BALB/c mice) and with two distinct Mtb strains (25, 134, 135). Non-castrated male mice exhibited significantly higher mortality rates and bacillary burdens compared with female and castrated male mice, supporting a role for testosterone in increased disease susceptibility (134). In addition, BCG-vaccinated female mice showed greater protection against TB than males in two mouse models (136, 137). Collectively, these results indicate that females develop superior protective immunity against Mtb compared with males. However, this is not to discount the contribution of non-biological factors to the increased risk of TB disease in human males, which is likely to be significant.
Males and females exhibit important sex-specific differences in the B cell compartment, including variations in B cell numbers, activation status, and antibody production. Females generally display higher B cell counts and mount stronger humoral immune responses, producing higher antibody levels following infection or vaccination (133). Consistent with this, recent studies suggest notable sex differences in the formation, structure, and function of iBALT. In COPD, the development of lymphoid follicles is more pronounced in female mice upon exposure to cigarette smoke (138). These differences were abolished with ovariectomy, suggesting that female sex hormones play a key role in lymphoid follicle development. While their exact role in COPD remains unclear, lymphoid follicles have been linked to severe disease progression (139). Notably, females appear to be more susceptible to developing COPD and exhibit increased expression of lymphoid follicles in the airways of COPD-affected lungs (138, 140). Similarly, in a model of collagen-induced arthritis (CIA), Dimitrijević et al. (2020) (141) observed sex differences in the generation of T follicular helper cells (Tfh), a subset of CD4+ T cells crucial for B cell responses and germinal center formation. Female rats, who are more susceptible to CIA, exhibited significantly higher frequencies of Tfh cells, resulting in stronger GC B cell formation, elevated antibody responses, and more severe disease progression compared to males (141). In contrast, Mtb infected female mice –who exhibit improved long-term control of Mtb infection compared to males– develop larger B cell follicles than their male counterparts (25). The smaller size of B cell follicles in males, which is associated with increased disease progression and higher mortality rates, underscores the protective role of B cell follicles in TB immunity. Several cytokines critical for the formation and maintenance of iBALT are upregulated in females compared to males across various diseases (Table 1). Importantly, cytokines essential for protective immunity in TB, such as IL-1α, IL-6, IL-23, and IL-17 (156–159) play a crucial role in the induction and formation of iBALT and are elevated in Mtb-infected females compared to males. More recently, Swanson et al. (2023) demonstrated a crucial role of antigen-specific B cells in the strategic localization of Tfh-like cells within iBALT to mediate Mtb control (40). According to Linge et al., iBALT structures in TB-susceptible mice were populated by B cells lacking TB antigen specificity, underscoring once again the critical role of antigen-specificity in mounting a protective iBALT-mediated immune response (160). However, the influence of the biological sex on the generation of Tfh cells and the close interaction with B cells in iBALT in TB has yet to be elucidated.
iBALT formationdiseasespeciesSexReferencesCytokinesCXCL13/
CXCR5TBMouseNo differences(26)CCL19/
CCL21/CCR7TBMouseIncreased in females(26)IL-7CIAMouseNo differences(141)IL-17CIA
TB
UTI
InfluenzaMouse
Mouse
Mouse
MouseIncreased in females(26, 141–143)IL-1αTBMouseIncreased in females26IL-6TB
SLE
Influenza
Influenza (vaccination)Mouse
Mouse
Mouse
HumanIncreased in females(143–146)IL-21CIA
SLEMouse
MouseIncreased in females(141, 147)Type I IFNSLE
HIV
COVID-19
Hanta (Seoul)Human
Human
Human
RatIncreased in females(148–151)IL-23TBMouseIncreased in females(26)IL-27CIAMouseIncreased in females(141)IL-2CIAMouseIncreased in males(141)BAFFCOPD
SLEMouse/Human
Mouse/HumanIncreased in females(138, 152)Cell typesB cells–HumanIncreased in females(133, 153, 154)CD4+ T cells–Mouse/HumanIncreased in females(133, 155)Tfh cellsCIA
SLEMouse
MouseIncreased in females(141, 144, 147)
Sex differences in iBALT formation.
TB, tuberculosis; CIA, collagen-induced arthritis; UTI, urinary tract infection; SLE, systemic lupus erythematosus; COPD, chronic obstructive pulmonary disease.
In summary, sex-specific differences in iBALT formation (Figure 3) contribute to the generally more robust immune responses in females, which may play an important role in the defense against infections like TB, but also in the higher prevalence of certain autoimmune and inflammatory diseases in the female population.
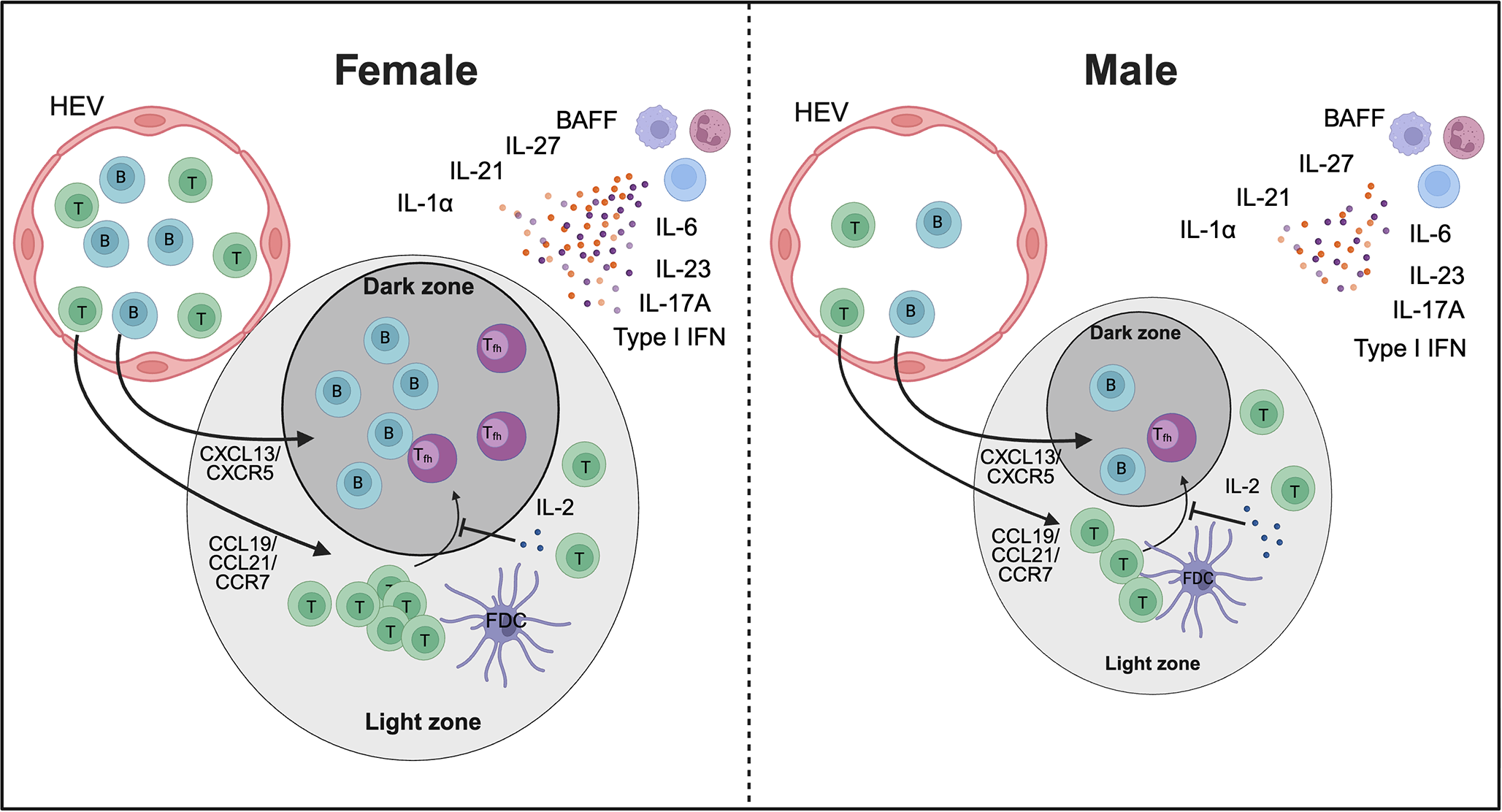
Proposed model of sex differences in iBALT formation. Higher numbers of B cells and T cells, particularly CD4+ T cells, along with increased levels of cytokines involved in iBALT development, may promote greater recruitment of these cells in females compared to males, potentially resulting in larger iBALT structures. In addition, elevated IL-21 and IL-27 together with reduced IL-2 could enhance Tfh cell differentiation in females. Together, these mechanisms may contribute to sex-specific differences in immunity, autoimmunity, and vaccine responses. HEV – high endothelial venules. The image was created using Biorender.
7 Discussion and concluding remarks
B cells play a far more dynamic and essential role in TB immunity than previously appreciated. Their involvement in antigen presentation, cytokine secretion, and TLS formation suggests that they are integral to shaping local immune responses in the lung. The discovery of iBALT as a potential site of protective immune activation challenges the conventional T cell-centric view of TB immunity and highlights new opportunities for immunotherapeutic strategies. The dual nature of iBALT—as both a protective structure and a contributor to pathology—necessitates a nuanced understanding of its regulation. Notably, IL-10-producing regulatory B cells may play a pivotal role in modulating TB immunity. While some B cell subsets contribute to protective responses, IL-10-secreting B cells could suppress pro-inflammatory immune activation, potentially limiting excessive lung damage but also creating an immunoregulatory environment that favors persistent Mtb infection. The balance between protective and immunosuppressive B cell functions within iBALT may be critical in determining TB disease outcomes.
Building on the evidence summarized in this review, we propose that B cells and iBALT could advance TB vaccine development and immunotherapy through several mechanisms. iBALT forms a lung-resident, germinal center-like niche where B cells undergo antigen-driven selection, class switching, and affinity maturation at the site of Mtb infection, producing high-affinity IgA and IgG that opsonize bacilli, neutralize secreted factors, and engage Fc-mediated effector pathways. B cells also act as antigen-presenting cells and cytokine producers, shaping local Th1, Th17, and Tfh responses and granuloma organization. Next-generation TB vaccines could be designed to induce protective iBALT using mucosal delivery platforms, adjuvants that promote lymphoid neogenesis and CXCL13-dependent recruitment, and antigens efficiently captured and presented by B cells, thereby establishing long-lived, tissue-resident B and T cell immunity in the lung. In parallel, defining B cell antigenic specificities, isotypes, and functional profiles in controlled versus progressive disease can guide selection of Mtb antigens and antibody subclasses for monoclonal therapies to enhance clearance, shorten chemotherapy, or prevent reactivation. This strategy may also identify targets for host-directed interventions that modulate B cell or iBALT function to reinforce protective rather than pathological lung immunity.
StatementsAuthor contributions
TSD: Conceptualization, Funding acquisition, Supervision, Visualization, Writing – original draft, Writing – review & editing. DH: Conceptualization, Resources, Visualization, Writing – original draft, Writing – review & editing. RK: Conceptualization, Resources, Visualization, Writing – original draft, Writing – review & editing. MH-T: Writing – review & editing. AL: Writing – review & editing. BS: Resources, Writing – review & editing.
Funding
The author(s) declared that financial support was received for this work and/or its publication. TSD acknowledges funding support from the National Institute of Allergy and Infectious Diseases (NIAID), NIH through the K99/R00 Pathway to Independence Award (1K99AI173401-01), which has been instrumental in supporting her research on B cell-mediated immunity in tuberculosis. RK is thankful for grant from the UK Medical Research Foundation (MRF-131-0002-RG-ELKIC0850) and Sub-Saharan African Network for TB/HIV Research Excellence (SANTHE) path to independence award #SANTHE-PTI007.
Acknowledgments
This review reflects a global collaboration among early career researchers from the USA, South Africa, and Germany. We are especially grateful to our mentors and colleagues who provided valuable insights during the development of this manuscript.
Conflict of interest
The author(s) declared that this work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declared that generative AI was not used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Global tuberculosis report (2025). Available online at: https://www.who.int/teams/global-programme-on-tuberculosis-and-lung-health/tb-reports/global-tuberculosis-report-2025 (Accessed December 11, 2025).
ZhuangLYeZLiLYangLGongW.
Next-generation TB vaccines: progress, challenges, and prospects. Vaccines. (2023) 11:1304. doi: 10.3390/vaccines11081304
SachdevaKSChadhaVK.
TB-vaccines: Current status & challenges. Indian J Med Res. (2024) 160:338–45. doi: 10.25259/IJMR_1478_2024
LarsenSEWilliamsBDRaisMColerRNBaldwinSL.
It takes a village: the multifaceted immune response to mycobacterium tuberculosis infection and vaccine-induced immunity. Front Immunol. (2022) 13:840225. doi: 10.3389/fimmu.2022.840225
NemesEFiore-GartlandABoggianoCCocciaMD’SouzaPGilbertPet al.
The quest for vaccine-induced immune correlates of protection against tuberculosis. Vaccine Insights. (2022) 1:165–81. doi: 10.18609/vac/2022.027
BeharSM.
Antigen-specific CD8+ T cells and protective immunity to tuberculosis. Adv Exp Med Biol. (2013) 783:141–63. doi: 10.1007/978-1-4614-6111-1_8
LinPLRutledgeTGreenAMBigbeeMFuhrmanCKleinEet al.
CD4 T cell depletion exacerbates acute Mycobacterium tuberculosis while reactivation of latent infection is dependent on severity of tissue depletion in cynomolgus macaques. AIDS Res Hum Retroviruses. (2012) 28:1693–702. doi: 10.1089/AID.2012.0028
MpandeCAMRozotVMositoBMusvosviMDintweOBBilekNet al.
Immune profiling of Mycobacterium tuberculosis-specific T cells in recent and remote infection. eBioMedicine. (2021) 64:103233. doi: 10.1016/j.ebiom.2021.103233
UrdahlKBShafianiSErnstJD.
Initiation and regulation of T-cell responses in tuberculosis. Mucosal Immunol. (2011) 4:288–93. doi: 10.1038/mi.2011.10
OgongoPPorterfieldJZLeslieA.
Lung tissue resident memory T-cells in the immune response to mycobacterium tuberculosis. Front Immunol. (2019) 10:992. doi: 10.3389/fimmu.2019.00992
Flores-GonzalezJUrbán-SolanoARamón-LuingLACancino-DiazJCContreras-RodriguezACuriel-QuesadaEet al.
Active tuberculosis patients have high systemic IgG levels and B cells fingerprinting, characterized by a reduced capacity to produce IFN-γ or IL-10 as a response to M.tb antigens. Front Immunol. (2023) 14:1263458. doi: 10.3389/fimmu.2023.1263458
DuttTSChoreño-ParraJA.
Editorial: Tuberculosis and humoral immunity. Front Immunol. (2025) 16:1562567. doi: 10.3389/fimmu.2025.1562567
WangQNagDBaldwinSLColerRNMcNamaraRP.
Antibodies as key mediators of protection against Mycobacterium tuberculosis. Front Immunol. (2024) 15:1430955. doi: 10.3389/fimmu.2024.1430955
KrauseROgongoPTezeraLAhmedMMbanoIChambersMet al.
B cell heterogeneity in human tuberculosis highlights compartment-specific phenotype and functional roles. Commun Biol. (2024) 7:1–15. doi: 10.1038/s42003-024-06282-7
DuttTSKargerBRFoxAYoussefNDadhwalRAliMZet al.
Mucosal exposure to non-tuberculous mycobacteria elicits B cell-mediated immunity against pulmonary tuberculosis. Cell Rep. (2022) 41:111783. doi: 10.1016/j.celrep.2022.111783
ChenYBharrhanSXuJSharmaTWangYSalgamePet al.
B cells promote granulomatous inflammation during chronic Mycobacterium tuberculosis infection in mice. PloS Pathog. (2023) 19:e1011187. doi: 10.1371/journal.ppat.1011187
BatistaFDHarwoodNE.
The who, how and where of antigen presentation to B cells. Nat Rev Immunol. (2009) 9:15–27. doi: 10.1038/nri2454
LundFE.
Cytokine-producing B lymphocytes – key regulators of immunity. Curr Opin Immunol. (2008) 20:332–8. doi: 10.1016/j.coi.2008.03.003
VazquezMICatalan-DibeneJZlotnikA.
B cells responses and cytokine production are regulated by their immune microenvironment. Cytokine. (2015) 74:318–26. doi: 10.1016/j.cyto.2015.02.007
DunlapMDPrinceOARangel-MorenoJThomasKAScordoJMTorrellesJBet al.
Formation of lung inducible bronchus associated lymphoid tissue is regulated by mycobacterium tuberculosis expressed determinants. Front Immunol. (2020) 11:1325. doi: 10.3389/fimmu.2020.01325
NagatakeTWadaYMatsumotoNShimojouMHirataSNasuAet al.
Inducible bronchus-associated lymphoid tissue plays an important role in the induction of antigen-specific immune response by Ag85B-hPIV2-based anti-tuberculosis vaccine in mice. J Immunol. (2016) 196:68. doi: 10.4049/jimmunol.196.Supp.68.9
KaushalDForemanTWGautamUSAlvarezXAdekambiTRangel-MorenoJet al.
Mucosal vaccination with attenuated Mycobacterium tuberculosis induces strong central memory responses and protects against tuberculosis. Nat Commun. (2015) 6:8533. doi: 10.1038/ncomms9533
MaglionePJXuJChanJ.
B Cells Moderate Inflammatory Progression and Enhance Bacterial Containment upon Pulmonary Challenge with Mycobacterium tuberculosis1. J Immunol. (2007) 178:7222–34. doi: 10.4049/jimmunol.178.11.7222
HertzDMarwitzSEggersLvon BorstelLHarikumar ParvathyGBehrendsJet al.
Sex-specific impact of B cell-derived IL-10 on tuberculosis resistance. Front Immunol. (2025) 16:1524500. doi: 10.3389/fimmu.2025.1524500
HertzDDibbernJEggersLvon BorstelLSchneiderBE.
Increased male susceptibility to Mycobacterium tuberculosis infection is associated with smaller B cell follicles in the lungs. Sci Rep. (2020) 10:5142. doi: 10.1038/s41598-020-61503-3
HertzDSchneiderB.
Sex differences in tuberculosis. Semin Immunopathol. (2019) 41:225–37. doi: 10.1007/s00281-018-0725-6
GuptaMSrikrishnaGKleinSBishaiWR.
Genetic and hormonal mechanisms underlying sex-specific immune responses in tuberculosis. Trends Immunol. (2022) 43:640–56. doi: 10.1016/j.it.2022.06.004
LeeRDMunroSAKnutsonTPLaRueRSHeltemes-HarrisLMFarrarMA.
Single-cell analysis identifies dynamic gene expression networks that govern B cell development and transformation. Nat Commun. (2021) 12:6843. doi: 10.1038/s41467-021-27232-5
HaoDHanGSinjabAGomez-BolanosLILazcanoRSerranoAet al.
The single-cell immunogenomic landscape of B and plasma cells in early-stage lung adenocarcinoma. Cancer Discov. (2022) 12:2626–45. doi: 10.1158/2159-8290.CD-21-1658
LuLLChungAWRosebrockTRGhebremichaelMYuWHGracePSet al.
A functional role for antibodies in tuberculosis. Cell. (2016) 167:433–443.e14. doi: 10.1016/j.cell.2016.08.072
KawaharaJYIrvineEBAlterG.
A case for antibodies as mechanistic correlates of immunity in tuberculosis. Front Immunol. (2019) 10:996. doi: 10.3389/fimmu.2019.00996
PhuahJYMattilaJTLinPLFlynnJL.
Activated B cells in the granulomas of nonhuman primates infected with Mycobacterium tuberculosis. Am J Pathol. (2012) 181:508–14. doi: 10.1016/j.ajpath.2012.05.009
SpatolaMNzizaNIrvineEBCizmeciDJungWVanLHet al.
Distinctive antibody responses to Mycobacterium tuberculosis in pulmonary and brain infection. Brain. (2024) 147:3247–60. doi: 10.1093/brain/awae066
JoostenSAvanMKEdelNFBaiocchiniAPetroneLVaniniVet al.
Patients with tuberculosis have a dysfunctional circulating B cells compartment, which normalizes following successful treatment. PloS Pathog. (2016) 12:e1005687. doi: 10.1371/journal.ppat.1005687
CorominasMCardonaVGonzalezLCaylàJARufiGMestreMet al.
B-lymphocytes and co-stimulatory molecules in Mycobacterium tuberculosis infection. Int J Tuberc Lung Dis Off J Int Union Tuberc Lung Dis. (2004) 8:98–105.
HernandezJVelazquezCValenzuelaORobles-ZepedaRRuiz-BustosENavarroMet al.
Low number of peripheral blood B lymphocytes in patients with pulmonary tuberculosis. Immunol Invest. (2010) 39:197–205. doi: 10.3109/08820130903586346
Roy ChowdhuryRVallaniaFYangQLopez AngelCJDarboeFPenn-NicholsonAet al.
A multi-cohort study of the immune factors associated with M. tuberculosis infection outcomes. Nat. (2018) 560:644–8. doi: 10.1038/s41586-018-0439-x
AlbertsBJohnsonALewisJRaffMRobertsKWalterP.
B cells and antibodies. In: Molecular biology of the cell (2002). New York, London:
Garland Science. Available online at: https://www.ncbi.nlm.nih.gov/books/NBK26884/. Garland Science.
HaasKM.
Non-canonical B cells: characteristics of uncharacteristic B cells. J Immunol Baltim Md. (2023) 1950:211. doi: 10.4049/jimmunol.2200944
SwansonRVGuptaAForemanTWLuLChoreno-ParraJAMbandiSKet al.
Antigen-specific B cells direct T follicular-like helper cells into lymphoid follicles to mediate Mycobacterium tuberculosis control. Nat Immunol. (2023) 24:855–68. doi: 10.1038/s41590-023-01476-3
AkkayaMKwakKPierceSK.
B cell memory: building two walls of protection against pathogens. Nat Rev Immunol. (2020) 20:229–38. doi: 10.1038/s41577-019-0244-2
OnoderaTTakahashiYYokoiYAtoMKodamaYHachimuraSet al.
Memory B cells in the lung participate in protective humoral immune responses to pulmonary influenza virus reinfection. Proc Natl Acad Sci. (2012) 109:2485–90. doi: 10.1073/pnas.1115369109
GermainCGnjaticSTamzalitFKnockaertSRemarkRGocJet al.
Presence of B cells in tertiary lymphoid structures is associated with a protective immunity in patients with lung cancer. Am J Respir Crit Care Med. (2014) 189:832–44. doi: 10.1164/rccm.201309-1611OC
HughesCEBensonRABedajMMaffiaP.
Antigen-presenting cells and antigen presentation in tertiary lymphoid organs. Front Immunol. (2016) 7:481. doi: 10.3389/fimmu.2016.00481
NaderiWSchreinerDKingCG.
T-cell-B cells collaboration in the lung. Curr Opin Immunol. (2023) 81:102284. doi: 10.1016/j.coi.2023.102284
LuLLSuscovichTJFortuneSMAlterG.
Beyond binding: antibody effector functions in infectious diseases. Nat Rev Immunol. (2018) 18:46–61. doi: 10.1038/nri.2017.106
IrvineEBNikolovAKhanMZPetersJMLuRSixsmithJet al.
Fc-engineered antibodies promote neutrophil-dependent control of Mycobacterium tuberculosis. Nat Microbiol. (2024) 9:2369–82. doi: 10.1038/s41564-024-01777-9
KangPBAzadAKTorrellesJBKaufmanTMBeharkaATibesarEet al.
The human macrophage mannose receptor directs Mycobacterium tuberculosis lipoarabinomannan-mediated phagosome biogenesis. J Exp Med. (2005) 202:987–99. doi: 10.1084/jem.20051239
KumarSKSinghPSinhaS.
Naturally produced opsonizing antibodies restrict the survival of Mycobacterium tuberculosis in human macrophages by augmenting phagosome maturation. Open Biol. (2015) 5:150171. doi: 10.1098/rsob.150171
Correia-NevesMSundlingCCooperAKälleniusG.
Lipoarabinomannan in active and passive protection against tuberculosis. Front Immunol. (2019) 10:1968. doi: 10.3389/fimmu.2019.01968
IshidaECorriganDTMalonisRJHofmannDChenTAminAGet al.
Monoclonal antibodies from humans with Mycobacterium tuberculosis exposure or latent infection recognize distinct arabinomannan epitopes. Commun Biol. (2021) 4:1–13. doi: 10.1038/s42003-021-02714-w
LuLLSmithMTYuKKQLuedemannCSuscovichTJGracePSet al.
IFN-γ-independent immune markers of Mycobacterium tuberculosis exposure. Nat Med. (2019) 25:977–87. doi: 10.1038/s41591-019-0441-3
MaglionePJXuJCasadevallAChanJ.
Fc gamma receptors regulate immune activation and susceptibility during Mycobacterium tuberculosis infection. J Immunol Baltim Md 1950. (2008) 180:3329–38. doi: 10.4049/jimmunol.180.5.3329
WilliamsAReljicRNaylorIClarkSOFalero-DiazGSinghMet al.
Passive protection with immunoglobulin A antibodies against tuberculous early infection of the lungs. Immunology. (2004) 111:328–33. doi: 10.1111/j.1365-2567.2004.01809.x
Glatman-FreedmanACasadevallA.
Serum Therapy for Tuberculosis Revisited: Reappraisal of the Role of Antibody-Mediated Immunity against Mycobacterium tuberculosis. Clin Microbiol Rev. (1998) 11:514–32. doi: 10.1128/CMR.11.3.514
GuiradoEAmatIGilODíazJArcosVCaceresNet al.
Passive serum therapy with polyclonal antibodies against Mycobacterium tuberculosis protects against post-chemotherapy relapse of tuberculosis infection in SCID mice. Microbes Infect. (2006) 8:1252–9. doi: 10.1016/j.micinf.2005.12.004
McLeanMRLuLLKentSJChungAW.
An inflammatory story: antibodies in tuberculosis comorbidities. Front Immunol. (2019) 10:2846. doi: 10.3389/fimmu.2019.02846
YuanCQuZ-LTangX-LLiuQLuoWHuangCet al.
Mycobacterium tuberculosis mannose-capped lipoarabinomannan induces IL-10-producing B cells and hinders CD4+Th1 immunity. iScience. (2019) 11:13–30. doi: 10.1016/j.isci.2018.11.039
Moreira-TeixeiraLRedfordPSStavropoulosEGhilardiNMaynardCLWeaverCTet al.
O’Garra A. T cell-derived IL-10 impairs host resistance to mycobacterium tuberculosis infection. J Immunol Baltim Md. (2017) 1950:199. doi: 10.4049/jimmunol.1601340
DaiY-CWangW-DZhangJ-AChenCLuoH-LXuHet al.
MTB driven B cells producing IL-35 and secreting high level of IL-10 in the patients with active pulmonary tuberculosis. Mol Immunol. (2019) 112:175–81. doi: 10.1016/j.molimm.2019.05.004
ChenCXuHPengYLuoHHuangG-XWuX-Jet al.
Elevation in the counts of IL-35-producing B cells infiltrating into lung tissue in mycobacterial infection is associated with the downregulation of Th1/Th17 and upregulation of Foxp3+Treg. Sci Rep. (2020) 10:13212. doi: 10.1038/s41598-020-69984-y
BénardASakwaISchierlohPColomAMercierITailleuxLet al.
B cells producing type I IFN modulate macrophage polarization in tuberculosis. Am J Respir Crit Care Med. (2018) 197:801–13. doi: 10.1164/rccm.201707-1475OC
TsaiC-YOoMPehJHYeoBCMAptekmannALeeBet al.
Splenic marginal zone B cells restrict Mycobacterium tuberculosis infection by shaping the cytokine pattern and cell-mediated immunity. Cell Rep. (2024) 43:114426. doi: 10.1016/j.celrep.2024.114426
LingeITsarevaAKondratievaEDyatlovAHidalgoJZvartsevRet al.
Pleiotropic effect of IL-6 produced by B-lymphocytes during early phases of adaptive immune responses against TB infection. Front Immunol. (2022) 13:750068. doi: 10.3389/fimmu.2022.750068
ChenYWuYYanGZhangG.
Tertiary lymphoid structures in cancer: maturation and induction. Front Immunol. (2024) 15:1369626. doi: 10.3389/fimmu.2024.1369626
ZhaoLJinSWangSZhangZWangXChenZet al.
Tertiary lymphoid structures in diseases: immune mechanisms and therapeutic advances. Signal Transduct Target Ther. (2024) 9:1–43. doi: 10.1038/s41392-024-01947-5
KhanalSWielandAGundersonAJ.
Mechanisms of tertiary lymphoid structure formation: cooperation between inflammation and antigenicity. Front Immunol. (2023) 14:1267654. doi: 10.3389/fimmu.2023.1267654
GaoXLiuCWangS.
Mucosal immune responses in the lung during respiratory infection: The organization and regulation of iBALT structure. hLife. (2023) 1:71–82. doi: 10.1016/j.hlife.2023.09.001
Inducible bronchus-associated lymphoid tissue: taming inflammation in the lung – pubMed. Available online at (Accessed December 11, 2025).
TanH-XEsterbauerRVandervenHAJunoJAKentSJWheatleyAK.
Inducible bronchus-associated lymphoid tissues (iBALT) serve as sites of B cell selection and maturation following influenza infection in mice. Front Immunol. (2019) 10:611. doi: 10.3389/fimmu.2019.00611
FooSYPhippsS.
Regulation of inducible BALT formation and contribution to immunity and pathology. Mucosal Immunol. (2010) 3:537–44. doi: 10.1038/mi.2010.52
EddensTElsegeinyWGarcia-Hernadez M de laLCastilloPTrevejo-NunezGSerodyKet al.
Pneumocystis-driven inducible bronchus-associated lymphoid tissue formation requires th2 and th17 immunity. . Cell Rep. (2017) 18:3078–90. doi: 10.1016/j.celrep.2017.03.016
Rangel-MorenoJCarragherDMde la Luz Garcia-HernandezMHwangJYKusserKHartsonLet al.
The development of inducible bronchus-associated lymphoid tissue depends on IL-17. Nat Immunol. (2011) 12:639–46. doi: 10.1038/ni.2053
Silva-SanchezARandallTD.
Role of iBALT in respiratory immunity. Curr Top Microbiol Immunol. (2020) 426:21–43. doi: 10.1007/82_2019_191
Rangel-MorenoJHartsonLNavarroCGaxiolaMSelmanMRandallTD.
Inducible bronchus-associated lymphoid tissue (iBALT) in patients with pulmonary complications of rheumatoid arthritis. J Clin Invest. (2006) 116:3183–94. doi: 10.1172/JCI28756
ZhaoRGaoD.
Innate immunity and tertiary lymphoid structures. Immunol Rev. (2025) 332:e70052. doi: 10.1111/imr.70052
NagatakeTSuzukiHHirataSMatsumotoNWadaYMorimotoSet al.
Immunological association of inducible bronchus-associated lymphoid tissue organogenesis in Ag85B-rHPIV2 vaccine-induced anti-tuberculosis mucosal immune responses in mice. Int Immunol. (2018) 30:471–81. doi: 10.1093/intimm/dxy046
FlynnRDuJVeenstraRGReichenbachDKPanoskaltsis-MortariATaylorPAet al.
Increased T follicular helper cells and germinal center B cells are required for cGVHD and bronchiolitis obliterans. Blood. (2014) 123:3988–98. doi: 10.1182/blood-2014-03-562231
McDonald-HymanCFlynnRPanoskaltsis-MortariAPetersonNMacDonaldKPAHillGRet al.
Therapeutic regulatory T-cell adoptive transfer ameliorates established murine chronic GVHD in a CXCR5-dependent manner. Blood. (2016) 128:1013–7. doi: 10.1182/blood-2016-05-715896
LiWGauthierJMHigashikuboRHsiaoH-MTanakaSVuongLet al.
Bronchus-associated lymphoid tissue–resident Foxp3+ T lymphocytes prevent antibody-mediated lung rejection. J Clin Invest. (2019) 129:556–68. doi: 10.1172/JCI122083
ZhangMWangZGranerMWYangLLiaoMYangQet al.
B cell infiltration is associated with the increased IL-17 and IL-22 expression in the lungs of patients with tuberculosis. Cell Immunol. (2011) 270:217–23. doi: 10.1016/j.cellimm.2011.05.009
Moyron-QuirozJERangel-MorenoJKusserKHartsonLSpragueFGoodrichSet al.
Role of inducible bronchus associated lymphoid tissue (iBALT) in respiratory immunity. Nat Med. (2004) 10:927–34. doi: 10.1038/nm1091
IwataMSatoA.
Morphological and immunohistochemical studies of the lungs and bronchus-associated lymphoid tissue in a rat model of chronic pulmonary infection with Pseudomonas aeruginosa. Infect Immun. (1991) 59:1514–20. doi: 10.1128/iai.59.4.1514-1520.1991
KitazawaHSatoAIwataM.
A study of bronchus-associated lymphoid tissue in a rat model of chronic pulmonary infection with pseudomonas aeruginosa. J Jpn Assoc Infect Dis. (1997) 71:214–21. doi: 10.11150/kansenshogakuzasshi1970.71.214
HalleSDujardinHCBakocevicNFleigeHDanzerHWillenzonSet al.
Induced bronchus-associated lymphoid tissue serves as a general priming site for T cells and is maintained by dendritic cells. J Exp Med. (2009) 206:2593–601. doi: 10.1084/jem.20091472
SwainSDMeissnerNHanSHarmsenA.
Pneumocystis infection in an immunocompetent host can promote collateral sensitization to respiratory antigens. Infect Immun. (2011) 79:1905–14. doi: 10.1128/iai.01273-10
WileyJAHarmsenAG.
Pneumocystis infection enhances antibody-mediated resistance to a subsequent influenza infection1. J Immunol. (2008) 180:5613–24. doi: 10.4049/jimmunol.180.8.5613
AdachiYOnoderaTYamadaYDaioRTsuijiMInoueTet al.
Distinct germinal center selection at local sites shapes memory B cell response to viral escape. J Exp Med. (2015) 212:1709–23. doi: 10.1084/jem.20142284
Rangel-MorenoJCarragherDMGarcia-Hernandez M de laLHwangJYKusserKHartsonLet al.
The development of inducible Bronchus Associated Lymphoid Tissue (iBALT) is dependent on IL-17. . Nat Immunol. (2011) 12:639–46. doi: 10.1038/ni.2053
WileyJARichertLESwainSDHarmsenABarnardDLRandallTDet al.
Inducible bronchus-associated lymphoid tissue elicited by a protein cage nanoparticle enhances protection in mice against diverse respiratory viruses. PloS One. (2009) 4:e7142. doi: 10.1371/journal.pone.0007142
Moyron-QuirozJERangel-MorenoJHartsonLKusserKTigheMPKlonowskiKDet al.
Persistence and responsiveness of immunologic memory in the absence of secondary lymphoid organs. Immunity. (2006) 25:643–54. doi: 10.1016/j.immuni.2006.08.022
HolbrookBCHaywardSLBlevinsLKKockNAycockTParksGDet al.
Nonhuman primate infants have an impaired respiratory but not systemic IgG antibody response following influenza virus infection. Virology. (2015) 476:124–33. doi: 10.1016/j.virol.2014.12.007
TamiyaYNakaiTSuzukiAMimakiSTsuchiharaKSatoKet al.
The impact of tertiary lymphoid structures on clinicopathological, genetic and gene expression characteristics in lung adenocarcinoma. Lung Cancer Amst Neth. (2022) 174:125–32. doi: 10.1016/j.lungcan.2022.11.001
KasikovaLRakovaJHenslerMLanickovaTTomankovaJPasulkaJet al.
Tertiary lymphoid structures and B cells determine clinically relevant T cell phenotypes in ovarian cancer. Nat Commun. (2024) 15:2528. doi: 10.1038/s41467-024-46873-w
RodriguezABPeskeJDWoodsANLeickKMMauldinISMeneveauMOet al.
Immune mechanisms orchestrate tertiary lymphoid structures in tumors via cancer-associated fibroblasts. Cell Rep. (2021) 36(3):109422. doi: 10.1016/j.celrep.2021.109422
SinghDKAhmedMAkterSShivannaVBucşanANMishraAet al.
Prevention of tuberculosis in cynomolgus macaques by an attenuated Mycobacterium tuberculosis vaccine candidate. Nat Commun. (2025) 16:1957. doi: 10.1038/s41467-025-57090-4
John-SchusterGGünterSHagerKConlonTMEickelbergOYildirimAÖ.
Inflammaging increases susceptibility to cigarette smoke-induced COPD. Oncotarget. (2016) 7:30068–83. doi: 10.18632/oncotarget.4027
ShinodaKHiraharaKNakayamaT.
Maintenance of pathogenic Th2 cells in allergic disorders. Allergol Int. (2017) 66:369–76. doi: 10.1016/j.alit.2017.03.005
PerrosFDorfmüllerPMontaniDHammadHWaelputWGirerdBet al.
Pulmonary lymphoid neogenesis in idiopathic pulmonary arterial hypertension. Am J Respir Crit Care Med. (2012) 185:311–21. doi: 10.1164/rccm.201105-0927OC
GopalRRangel-MorenoJJuneckoBAFMallonDJChenKPociaskDAet al.
Mucosal pre-exposure to th17-inducing adjuvants exacerbates pathology after influenza infection. Am J Pathol. (2014) 184:55–63. doi: 10.1016/j.ajpath.2013.09.012
GosmanMMEWillemseBWMJansenDFLapperreTSvanSAPSHet al.
Group the G and LUC in OLD (GLUCOLD) S. Increased number of B cellss in bronchial biopsies in COPD. Eur Respir J. (2006) 27:60–4. doi: 10.1183/09031936.06.00007005
ColvinKLCripePJIvyDDStenmarkKRYeagerME.
Bronchus-associated lymphoid tissue in pulmonary hypertension produces pathologic autoantibodies. Am J Respir Crit Care Med. (2013) 188:1126–36. doi: 10.1164/rccm.201302-0403OC
AuaisAAdkinsBNapchanGPiedimonteG.
Immunomodulatory effects of sensory nerves during respiratory syncytial virus infection in rats. Am J Physiol-Lung Cell Mol Physiol. (2003) 285:L105–13. doi: 10.1152/ajplung.00004.2003
JohnsonJEGonzalesRAOlsonSJWrightPFGrahamBS.
The histopathology of fatal untreated human respiratory syncytial virus infection. Mod Pathol. (2007) 20:108–19. doi: 10.1038/modpathol.3800725
HoltPGSlyPD.
Th2 cytokines in the asthma late-phase response. Lancet. (2007) 370:1396–8. doi: 10.1016/S0140-6736(07)61587-6
RobinsonPJHegeleRGSchellenbergRR.
Allergic sensitization increases airway reactivity in Guinea pigs with respiratory syncytial virus bronchiolitis. J Allergy Clin Immunol. (1997) 100:492–8. doi: 10.1016/S0091-6749(97)70141-6
MesinLErschingJVictoraGD.
Germinal center B cell dynamics. Immunity. (2016) 45:471–82. doi: 10.1016/j.immuni.2016.09.001
MaglionePJChanJ.
How B cells shape the immune response against Mycobacterium tuberculosis. Eur J Immunol. (2009) 39:676–86. doi: 10.1002/eji.200839148
WoodruffMCRamonellRPNguyenDCCashmanKSSainiASHaddadNSet al.
Extrafollicular B cell responses correlate with neutralizing antibodies and morbidity in COVID-19. Nat Immunol. (2020) 21:1506–16. doi: 10.1038/s41590-020-00814-z
MenonMHussellTAli ShuwaH.
Regulatory B cells in respiratory health and diseases. Immunol Rev. (2021) 299:61–73. doi: 10.1111/imr.12941
MauriCMenonM.
Human regulatory B cells in health and disease: therapeutic potential. J Clin Invest. (2017) 127:772–9. doi: 10.1172/JCI85113
TeoATanHDLoyTChiaPYChuaCLL.
Understanding antibody-dependent enhancement in dengue: Are afucosylated IgG1s a concern? PloS Pathog. (2023) 19:e1011223. doi: 10.1371/journal.ppat.1011223
ChanJMehtaSBharrhanSChenYAchkarJMCasadevallAet al.
The role of B cells and humoral immunity in Mycobacterium tuberculosis infection. Semin Immunol. (2014) 26:588–600. doi: 10.1016/j.smim.2014.10.005
KozakiewiczLPhuahJFlynnJChanJ.
The role of B cells and humoral immunity in mycobacterium tuberculosis infection.Adv Exp Med Biol. (2013) 783:225–50. doi: 10.1007/978-1-4614-6111-1_12
KahnertAHöpkenUESteinMBandermannSLippMKaufmannSHE.
Mycobacterium tuberculosis triggers formation of lymphoid structure in murine lungs. J Infect Dis. (2007) 195:46–54. doi: 10.1086/508894
Peralta AlvarezMPJonesHRedondo AzemaHDavisCWhiteADSarfasCet al.
Low-dose M.tb infection but not BCG or MTBVAC vaccination enhances heterologous antibody titres in non-human primates. Front Immunol. (2024) 15:1387454. doi: 10.3389/fimmu.2024.1387454
KimudaSGAndia-BiraroISebinaIEgesaMNalwogaASmithSGet al.
Mycobacterium tuberculosis infection is associated with increased B cell responses to unrelated pathogens. Sci Rep. (2020) 10:14324. doi: 10.1038/s41598-020-71044-4
OhmenJDBarnesPFGrissoCLBloomBRModlinRL.
Evidence for a superantigen in human tuberculosis. Immunity. (1994) 1:35–43. doi: 10.1016/1074-7613(94)90007-8
LevinsonAIKozlowskiLZhengYWheatleyL.
B cells superantigens: definition and potential impact on the immune response. J Clin Immunol. (1995) 15:26S–36S. doi: 10.1007/BF01540891
BernasconiNLTraggiaiELanzavecchiaA.
Maintenance of serological memory by polyclonal activation of human memory B cells. Science. (2002) 298:2199–202. doi: 10.1126/science.1076071
UlrichsTKosmiadiGAJörgSPradlLTitukhinaMMishenkoVet al.
Differential organization of the local immune response in patients with active cavitary tuberculosis or with nonprogressive tuberculoma. J Infect Dis. (2005) 192:89–97. doi: 10.1086/430621
UlrichsTKosmiadiGATrusovVJörgSPradlLTitukhinaMet al.
Human tuberculous granulomas induce peripheral lymphoid follicle-like structures to orchestrate local host defence in the lung. J Pathol. (2004) 204:217–28. doi: 10.1002/path.1628
PerdomoCZedlerUKühlAALozzaLSaikaliPSanderLEet al.
Mucosal BCG vaccination induces protective lung-resident memory T cell populations against tuberculosis. mBio. (2016) 7:10. doi: 10.1128/mbio.01686-16
AguiloNAlvarez-ArguedasSUrangaSMarinovaDMonzónMBadiolaJet al.
Pulmonary but not subcutaneous delivery of BCG vaccine confers protection to tuberculosis-susceptible mice by an interleukin 17–dependent mechanism. J Infect Dis. (2016) 213:831–9. doi: 10.1093/infdis/jiv503
GopalRRangel-MorenoJSlightSLinYNawarHFJuneckoBAFet al.
Interleukin-17-dependent CXCL13 mediates mucosal vaccine–induced immunity against tuberculosis. Mucosal Immunol. (2013) 6:972–84. doi: 10.1038/mi.2012.135
GrayJOehrleKWorthenGAlenghatTWhitsettJDeshmukhH.
Intestinal commensal bacteria mediate lung mucosal immunity and promote resistance of newborn mice to infection. Sci Transl Med. (2017) 9:eaaf9412. doi: 10.1126/scitranslmed.aaf9412
ArdainADomingo-GonzalezRDasSKazerSWHowardNCSinghAet al.
Group 3 innate lymphoid cells mediate early protective immunity against tuberculosis. Nature. (2019) 570:528–32. doi: 10.1038/s41586-019-1276-2
GriffithsKLAhmedMDasSGopalRHorneWConnellTDet al.
Targeting dendritic cells to accelerate T-cell activation overcomes a bottleneck in tuberculosis vaccine efficacy. Nat Commun. (2016) 7:13894. doi: 10.1038/ncomms13894
KaushalDMehraS.
Didier P j., Lackner A a. The non-human primate model of tuberculosis. J Med Primatol. (2012) 41:191–201. doi: 10.1111/j.1600-0684.2012.00536.x
DarrahPAZeppaJJMaielloPHackneyJAWadsworthMHHughesTKet al.
Prevention of tuberculosis in macaques after intravenous BCG immunization. Nature. (2020) 577:95–102. doi: 10.1038/s41586-019-1817-8
IrvineEBO’NeilADarrahPAShinSChoudharyALiWet al.
Robust IgM responses following intravenous vaccination with Bacille Calmette–Guérin associate with prevention of Mycobacterium tuberculosis infection in macaques. Nat Immunol. (2021) 22:1515–23. doi: 10.1038/s41590-021-01066-1
SimonsonAWZeppaJJBucsanANChaoMCPokkaliSHopkinsFet al.
Intravenous BCG-mediated protection against tuberculosis requires CD4+ T cells and CD8α+ lymphocytes. J Exp Med. (2025) 222:e20241571. doi: 10.1084/jem.20241571
KleinSLFlanaganKL.
Sex differences in immune responses. Nat Rev Immunol. (2016) 16:626–38. doi: 10.1038/nri.2016.90
BiniEIEspinosaDMCastilloBMPayánJBColucciDCruzAFet al.
The influence of sex steroid hormones in the immunopathology of experimental pulmonary tuberculosis. PloS One. (2014) 9:e93831. doi: 10.1371/journal.pone.0093831
DibbernJEggersLSchneiderBE.
Sex differences in the C57BL/6 model of Mycobacterium tuberculosis infection. Sci Rep. (2017) 7, 10957. doi: 10.1038/s41598-017-11438-z
NieuwenhuizenNEZylaJZedlerUBandermannSAbu AbedUBrinkmannVet al.
Weaker protection against tuberculosis in BCG-vaccinated male 129 S2 mice compared to females. Vaccine. (2021) 39:7253–64. doi: 10.1016/j.vaccine.2021.09.039
ParvathyGHHertzDBhandiwadDEggersLvon BorstelLBehrendsJet al.
Sex differences in vaccine-induced immunity and protection against mycobacterium tuberculosis. The Journal of Infectious Diseases. (2025) 232(5):1187–1197. doi: 10.1093/infdis/jiaf277
TamATanabeNChurgAWrightJLHoggJCSinDD.
Sex differences in lymphoid follicles in COPD airways. Respir Res. (2020) 21:46. doi: 10.1186/s12931-020-1311-8
HoggJCChuFUtokaparchSWoodsRElliottWMBuzatuLet al.
The nature of small-airway obstruction in chronic obstructive pulmonary disease. N Engl J Med. (2004) 350:2645–53. doi: 10.1056/NEJMoa032158
BarnesPJ.
Sex differences in chronic obstructive pulmonary disease mechanisms. Am J Respir Crit Care Med. (2016) 193:813–4. doi: 10.1164/rccm.201512-2379ED
DimitrijevićMArsenović-RaninNKosecDBufanBNacka-AleksićMPilipovićIet al.
Sex differences in Tfh cell help to B cells contribute to sexual dimorphism in severity of rat collagen-induced arthritis. Sci Rep. (2020) 10:1214. doi: 10.1038/s41598-020-58127-y
Zychlinsky ScharffARousseauMLacerda MarianoLCantonTConsiglioCRAlbertMLet al.
Sex differences in IL-17 contribute to chronicity in male versus female urinary tract infection. JCI Insight. (2019) 5:e122998. doi: 10.1172/jci.insight.122998
HoffmannJOtteAThieleSLotterHShuYGabrielG.
Sex differences in H7N9 influenza A virus pathogenesis. Vaccine. (2015) 33:6949–54. doi: 10.1016/j.vaccine.2015.08.044
KimSJZouYRGoldsteinJReizisBDiamondB.
Tolerogenic function of Blimp-1 in dendritic cells. J Exp Med. (2011) 208:2193–9. doi: 10.1084/jem.20110658
RobinsonDPLorenzoMEJianWKleinSL.
Elevated 17β-estradiol protects females from influenza A virus pathogenesis by suppressing inflammatory responses. PloS Pathog. (2011) 7:e1002149. doi: 10.1371/journal.ppat.1002149
PotluriTFinkALSylviaKEDhakalSVermillionMSVom SteegLet al.
Age-associated changes in the impact of sex steroids on influenza vaccine responses in males and females. NPJ Vaccines. (2019) 4:29. doi: 10.1038/s41541-019-0124-6
FosterADHaasMPuliaevaISoloviovaKPuliaevRViaCS.
Donor CD8 T cell activation is critical for greater renal disease severity in female chronic graft-vs.-host mice and is associated with increased splenic ICOS(hi) host CD4 T cells and IL-21 expression. Clin Immunol Orlando Fla. (2010) 136:61–73. doi: 10.1016/j.clim.2010.01.005
BerghöferBFrommerTHaleyGFinkLBeinGHacksteinH.
TLR7 ligands induce higher IFN-alpha production in females. J Immunol Baltim Md. (2006) 1950:177. doi: 10.4049/jimmunol.177.4.2088
MeierAChangJJChanESPollardRBSidhuHKKulkarniSet al.
Sex differences in the Toll-like receptor-mediated response of plasmacytoid dendritic cells to HIV-1. Nat Med. (2009) 15:955–9. doi: 10.1038/nm.2004
SpieringAEde VriesTJ.
Why females do better: the X chromosomal TLR7 gene-dose effect in COVID-19. Front Immunol. (2021) 12:756262. doi: 10.3389/fimmu.2021.756262
HannahMFBajicVBKleinSL.
Sex differences in the recognition of and innate antiviral responses to Seoul virus in Norway rats. Brain Behav Immun. (2008) 22:503–16. doi: 10.1016/j.bbi.2007.10.005
PanchanathanRChoubeyD.
Murine BAFF expression is up-regulated by estrogen and interferons: implications for sex bias in the development of autoimmunity. Mol Immunol. (2013) 53:15–23. doi: 10.1016/j.molimm.2012.06.013
AbdullahMChaiP-SChongM-YTohitERMRamasamyRPeiCPet al.
Gender effect on in vitro lymphocyte subset levels of healthy individuals. Cell Immunol. (2012) 272:214–9. doi: 10.1016/j.cellimm.2011.10.009
DoddKCMenonM.
Sex bias in lymphocytes: Implications for autoimmune diseases. Front Immunol. (2022) 13:945762. doi: 10.3389/fimmu.2022.945762
DunnSEPerryWAKleinSL.
Mechanisms and consequences of sex differences in immune responses. Nat Rev Nephrol. (2024) 20:37–55. doi: 10.1038/s41581-023-00787-w
Mayer-BarberKDAndradeBBBarberDLHienySFengCGCasparPet al.
Innate and adaptive interferons suppress IL-1α and IL-1β production by distinct pulmonary myeloid subsets during Mycobacterium tuberculosis infection. Immunity. (2011) 35:1023–34. doi: 10.1016/j.immuni.2011.12.002
MartinezANMehraSKaushalD.
Role of interleukin 6 in innate immunity to Mycobacterium tuberculosis infection. J Infect Dis. (2013) 207:1253–61. doi: 10.1093/infdis/jit037
KhaderSAGuglaniLRangel-MorenoJGopalRJuneckoBAFFountainJJet al.
IL-23 is required for long-term control of Mycobacterium tuberculosis and B cell follicle formation in the infected lung. J Immunol Baltim Md. (2011) 1950:187. doi: 10.4049/jimmunol.1101377
Okamoto YoshidaYUmemuraMYahagiAO’BrienRLIkutaKKishiharaKet al.
Essential role of IL-17A in the formation of a mycobacterial infection-induced granuloma in the lung. J Immunol Baltim Md. (2010) 1950:184. doi: 10.4049/jimmunol.0903332
LingeIDyatlovAKondratievaEAvdienkoVAptAKondratievaT.
B-lymphocytes forming follicle-like structures in the lung tissue of tuberculosis-infected mice: Dynamics, phenotypes and functional activity. Tuberculosis. (2017) 102:16–23. doi: 10.1016/j.tube.2016.11.005
Summary
Keywords
antibodies, B cells, iBALT, sex differences, TLS, tuberculosis
Citation
Dutt TS, Krause R, Hertz D, Henao-Tamayo M, Leslie A and Schneider B (2026) B cells and iBALT in TB immunity & pathogenesis. Front. Immunol. 17:1743572. doi: 10.3389/fimmu.2026.1743572
Received
10 November 2025
Revised
13 January 2026
Accepted
13 January 2026
Published
04 February 2026
Volume
17 – 2026
Edited by
Sudhir Paul, University of Texas Health Science Center at Houston, United States
Reviewed by
Bo Yan, Fudan University, China
Irina Linge, Central Tuberculosis Research Institute (RAMS), Russia
Updates

Check for updates
Copyright
© 2026 Dutt, Krause, Hertz, Henao-Tamayo, Leslie and Schneider.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alasdair Leslie, al.leslie@ahri.org; Marcela Henao-Tamayo, marcela.henao_tamayo@colostate.edu; Bianca Schneider, bschneider@fz-borstel.de
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share senior authorship
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.





