

Vitamin B12 induces the predatory morph and transgenerational memory
Long-term environmental induction is performed by switching naïve P. pacificus RSC011 cultures that were always grown on the standard laboratory food source E. coli OP50 to Novosphingobium L76 (Fig. 1b). This P. pacificus strain from La Réunion Island is preferentially non-predatory (30%Eu:70%St) when grown on E. coli. The original long-term environmental induction experiment with 110 isogenic lines derived from the same ancestral RSC011 individual revealed dietary induction of the predatory mouth form (Fig. 1b). This dietary effect is (i) immediate (in generation 1 on Novosphingobium), (ii) complete (100% Eu progeny), (iii) systemic (in all 110 lines), and (iv) permanent (throughout the entire experiment of 101 generations). Following the reversal from Novosphingobium back to the standard E. coli OP50 diet, we observed TEI of the predatory mouth form (Fig. 1b). Importantly, such TEI is only seen after a minimal exposure to Novosphingobium for five generations, whereas a shorter exposure will not result in memory of the predatory morph.
We wanted to determine the molecular nature of the dietary stimulus inducing the predatory mouth form and its transgenerational inheritance in P. pacificus RSC011. The Novosphingobium L76 strain used in this experimental setup was originally isolated from a Pristionchus-associated environment and was shown to enhance killing efficiency of the P. pacificus wild type strain PS312, which is naturally 100% Eu15. This study also identified bacterial vitamin B12 production to cause enhanced predation, to accelerate worm development and increase brood size15. Therefore, we wanted to know if vitamin B12 would also play a role in dietary induction of the predatory mouth form and TEI in P. pacificus RSC011 animals. For that, we transferred single naïve J4 animals to vitamin B12-supplemented agar plates with E. coli OP50 as food source (Fig. 2a). It is important to note that E. coli OP50 does not produce vitamin B12 but rather obtains its vitamin B12 from the tryptone in the agar15,16,17. We originally used final concentrations of 500, 1000, and 1500 nM of one of the two active forms of vitamin B12, methyl-Cobalamin (Me-Cbl), for supplementation, similar to previous studies in P. pacificus15. Note that these concentrations are substantially higher than those used in previous work in C. elegans17. Strikingly, we observed an immediate induction of the Eu mouth form after vitamin B12 supplementation at all three concentrations used (Fig. 2b–d and Supplementary Fig. 1b–d). However, vitamin B12 supplementation results in a strong but not a complete induction of the Eu mouth form. Specifically, in three independent biological replicates with a total of 60 assay plates, we found high but incomplete induction in the first generation after vitamin B12 supplementation (0.878 ≤ HDI (θ) ≤ 0.994, where θ is the inferred probability of developing the predatory mouth form). This effect was seen over a period of 5 and 10 generations and was observed in all lines (Fig. 2b–d and Supplementary Fig. 1b–d). Thus, vitamin B12 mimics Novosphingobium-based induction of the predatory mouth form in P. pacificus.
Fig. 2: Diet-derived vitamin B12 induces the predatory mouth form and its transgenerational inheritance.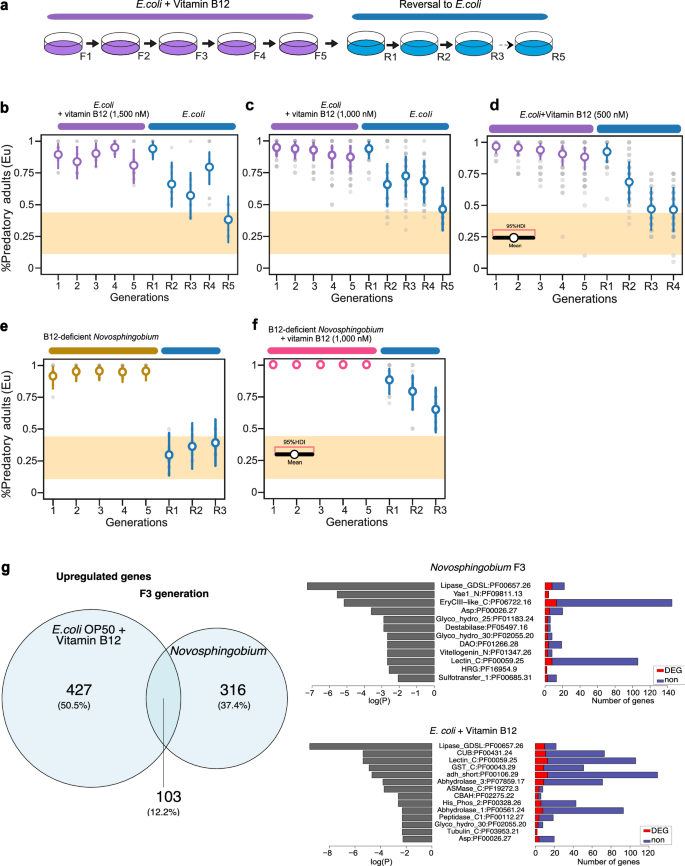
a Schematic diagram for vitamin B12 supplementation showing five generations on vitamin B12-supplemented plates and subsequent exposure to non-supplemented E. coli plates. b–d Vitamin B12 supplementation mimics induction and subsequent transgenerational inheritance of the Eu mouth form. Mean probability of predatory mouth form for five generations on 1500, 1000, and 500 nM vitamin B12-supplemented E. coli and five generations on non-supplemented E. coli. e Novosphingobium vitamin B12-deficient mutant induces the predatory morph but not its transgenerational inheritance. Mean probability of the predatory mouth form on a Novosphingobium vitamin B12-deficient mutant and reversal to E. coli. f Rescue of transgenerational inheritance of the predatory mouth form by supplementing exogenous vitamin B12 to Novosphingobium vitamin B12-deficient mutant. Mean probability of predatory mouth form for 5 generations on vitamin B12-supplemented Novosphingobium mutants and subsequent reversal to E. coli. Final mouth-form frequencies are the mean of at least 10 biological replicates (n = 20 animals per plate). Points represent the mean probability of developing the Eu morph, with error bars reflecting the 95% HDI from the Bayesian model. The yellow area indicates the RSC011 baseline response on E. coli, averaged over 101 generations from a previous study3. g Differential gene expression and enrichment analyses through single-worm transcriptomics (SWT). Exposure to Novosphingobium and vitamin B12-supplemented E. coli revealed overlapping differentially-expressed transcripts relative to worm cultures grown on a standard E. coli diet. The pathways with most significant enrichment (FDR-corrected p value < 0.01) for three generations on Novosphingobium and B12-supplemented E. coli are shown. SWT was performed using five independent biological replicates of young adult individuals. Naïve animals on E. coli were used as reference for differential expression analysis. See also Supplementary Fig. 1. Source data are provided as a Source Data file.
To determine if vitamin B12 would also induce TEI of the predatory mouth form, we established a mini-assay with a 5-generation or 10-generation exposure of worms to a vitamin B12-supplemented E. coli diet, followed by 5 generations on an un-supplemented diet (Fig. 2a–d and Supplementary Fig. 1a–d). Indeed, we observed TEI of the predatory mouth form (Fig. 2b–d and Supplementary Fig. 1b–d). Animals supplemented with 1000 and 1500 nM vitamin B12 showed TEI of the Eu mouth form similar to animals that were exposed to Novosphingobium for 5 or 10 generations (Fig. 2b, c and Supplementary Fig. 1b, c). In contrast, a supplementation with 500 nM of vitamin B12 only caused the induction of the predatory mouth form but no full TEI of the Eu morph (Fig. 2d). Specifically, the response during reversal only lasted for two generations, with all following generations being down to baseline Eu frequencies (0.841 ≤ HD1(θR1) ≤ 0.987, 0.52 ≤ HD1(θR2) ≤ 0.842, and 0.305 ≤ HD1(θR3) ≤ 0.637) (Fig. 2d and Supplementary Fig. 1d). Such a response is referred to as “intergenerational”, but not transgenerational inheritance4. These results allow two major conclusions. First, vitamin B12 supplementation mimics diet-derived induction of the predatory mouth form and its subsequent transgenerational inheritance. Second, the effect of vitamin B12 is concentration-dependent with only 1500 and 1000 nM but not 500 nM of vitamin B12 causing TEI.
Vitamin B12 induces a distinct transcriptomic response
Next, we wanted to know whether vitamin B12 supplementation can elicit similar effects on gene expression as those caused by the Novosphingobium diet. In principle, the Novosphingobium diet and vitamin B12 supplementation might induce similar transcriptomic responses or alternatively, have limited overlap in individual genes only. We examined the overlap of differential gene expression (DGE) found between vitamin B12 supplementation on E. coli OP50 with Novosphingobium-induced gene expression changes in RSC011 animals by using single worm transcriptomics of day 1 adult individuals (Fig. 2g). After three generations of exposure, Novosphingobium-upregulated genes were in parts also upregulated upon vitamin B12 supplementation (Fig. 2g and Supplementary Data 1). Specifically, of the 530 upregulated genes after a three-generation vitamin B12 supplementation, 103 (12.2% of all uniquely upregulated genes) were also upregulated on Novosphingobium. Notably, these shared genes correspond to 19.4% of the vitamin B12-induced and 24.6% of the Novosphingobium-induced transcriptional responses. Analysis of PFAM enrichment among these shared genes revealed that Lipase GDSL (PF00657.26) and Lectin_C (PF00059.25) were enriched in both conditions, suggesting that lipid metabolism and carbohydrate-binding/cell-surface signaling form a core transcriptional response. Beyond this overlap, vitamin B12-specific genes were uniquely enriched for CUB (PF00431.24), associated with extracellular protein interactions, whereas Novosphingobium-specific genes were enriched for Yae1_N (PF09811.13) and EryCIII-like_C (PF06722.16), reflecting ribosome-associated functions and secondary metabolite biosynthesis, respectively. These results highlight both conserved and condition-specific molecular programs activated in response to vitamin B12 supplementation and Novosphingobium exposure. (Fig. 2g and Supplementary Data 1). Thus, vitamin B12 supplementation to the E. coli OP50 diet exhibits limited overlap with Novosphingobium-induced gene expression changes.
Vitamin B12 is necessary and sufficient to induce memory
Previous studies that established a role of dietary vitamin B12 on life history traits in P. pacificus and C. elegans were based on screening Tn5-based bacterial mutant libraries15,17. We used a Novosphingobium vitamin B12-deficient mutant to study the effect of Novosphingobium-derived vitamin B12 on mouth-form plasticity and TEI of the predatory morph. Using the 5-generation mini-assay described above, we found that a Novosphingobium vitamin B12-deficient diet would still induce the Eu mouth form (Fig. 2e). However, in contrast to a wild-type Novosphingobium diet, the induction is not complete and plateaus ~ 0.95% Eu (0.818 ≤ HDI (θF1) ≤ 0.98 and 0.883 ≤ HDI(θF5) ≤ 0.999). These results indicate that additional factor(s) besides vitamin B12 are able to induce the predatory mouth form. Thus, the complete (100%) induction of the Eu morph of P. pacificus RSC011 on Novosphingobium is due to multiple factors.
In contrary, we found that the Novosphingobium vitamin B12-deficient diet would not cause TEI of the predatory mouth form. When we reverted worm lines that were exposed to the Novosphingobium vitamin B12-deficient diet for 5 generations back to E. coli, we observed an immediate return to the base line response level (0.135 ≤ HDI(θR1) ≤ 0.47) (Fig. 2e). Similarly, when we reverted RSC011 lines that had been exposed to this diet for 10 generations, we also did not observe TEI of the predatory mouth form (Supplementary Fig. 1e). In contrast, vitamin B12 supplementation for 5 or 10 generations to the Novosphingobium vitamin B12-deficient diet restored the normal TEI response after reversal to E. coli OP50 (0.772 ≤ HDI(θR1) ≤ 0.971, 0.65 ≤ HDI(θR2) ≤ 0.919, and 0.47 ≤HDI(θR3) ≤ 0.832) (Fig. 2f and Supplementary Fig. 1f). These results indicate that vitamin B12 is both necessary and sufficient to induce the TEI of the predatory mouth form.
Vitamin B12-induced TEI of the predatory morph requires metr-1
In animals, including humans, vitamin B12 acts as a cofactor of two enzymes, methionine-synthase in the cytoplasm and methylmalonyl coenzyme A (CoA) mutase in mitochondria (Fig. 3a)18. In P. pacificus as in C. elegans, the methionine-synthase is encoded by Ppa-metr-1. To manipulate the methylmalonyl coenzyme A (CoA) mutase pathway, we targeted the upstream factor Ppa-mce-1, that is one-to-one orthologous to Cel-mce-1. We targeted Ppa-metr-1 and Ppa-mce-1 in P. pacificus RSC011 by CRISPR to study a potential role in dietary induction and TEI of the predatory mouth form (Fig. 3b). We obtained two frameshift alleles of Ppa-metr-1 and Ppa-mce-1 each, and tested all of them in our standard assay (Fig. 3b). First, we studied the response of Ppa-metr-1 mutants to an E. coli diet supplemented with 500, 1000 and 1500 nM vitamin B12. Ppa-metr-1 mutant animals failed to respond to vitamin B12 indicating that the role of vitamin B12 in inducing the Eu mouth form requires Ppa-metr-1 (Fig. 3c). Second, we grew Ppa-metr-1 mutants on Novosphingobium and observed a mean induction of \(\bar{{{{\rm{\theta }}}}}\) = 0.96 throughout the exposure of 5–10 generations (Fig. 3d and Supplementary Fig. 2a, b). This result is likely due to the other Novosphingobium factor(s) that can induce the Eu mouth form as indicated in the experiments using the Novosphingobium vitamin B12-deficient diet (Fig. 2e and Supplementary Fig. 2e). Importantly, when we reverted Ppa-metr-1 mutants from Novosphingobium back to E. coli, we observed no TEI of the predatory mouth form (Fig. 3d and Supplementary Fig. 2a, b). These findings indicate that vitamin B12 acts as a cofactor of methionine-synthase in the transgenerational inheritance of the Eu mouth form. Third, we tested Ppa-mce-1 mutants and observed an induction of the Eu mouth form on Novosphingobium similar to wild-type animals (Fig. 3e and Supplementary Fig. 2d, e). After reversal, Ppa-mce-1 mutant animals responded similar to wild type controls, indicating that Ppa-MCE-1 is not involved in the TEI of the predatory mouth form (Fig. 3e and Supplementary Fig. 2d, e). Note however, that the response of Ppa-mce-1 mutants to vitamin B12-supplemented E. coli diet only resulted in a partial Eu induction (0.256 ≤ HDI (θ) ≤ 0.616 for the highest vitamin B12 concentration) (Supplementary Fig. 2c). Together, our mutant analysis identifies a requirement of Ppa-metr-1 for the vitamin B12-mediated induction of the predatory mouth form and its subsequent TEI.
Fig. 3: Vitamin B12 functions as a cofactor for methionine synthase metr-1 in the transgenerational inheritance of the Eu morph.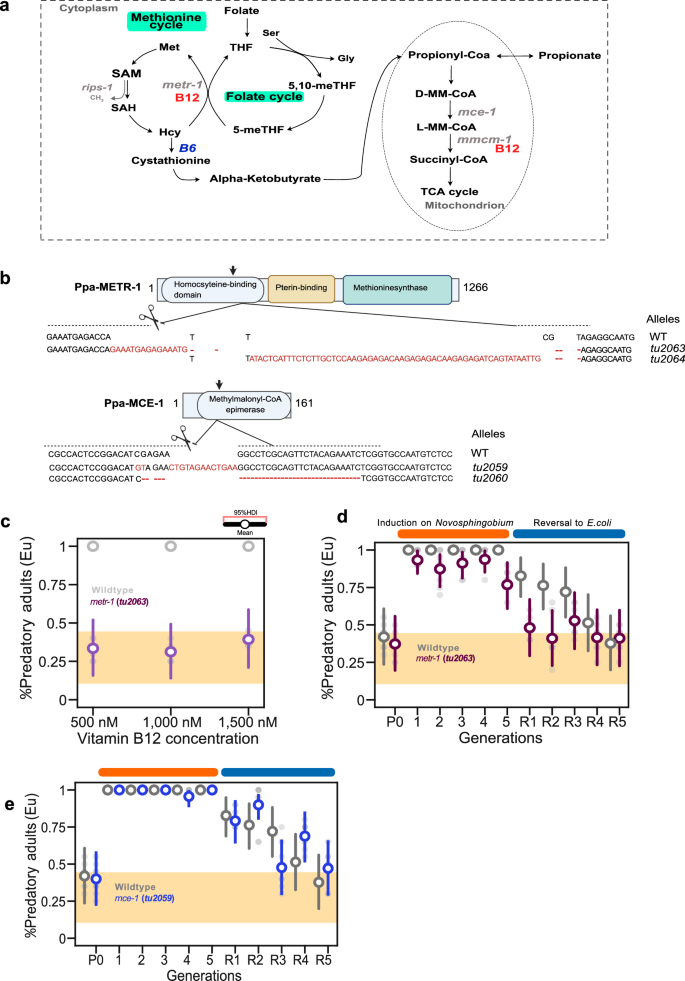
a Requirement for vitamin B12 as cofactor for two enzymes in the cytosol and mitochondria. In the cytoplasm, it acts as cofactor of methionine synthase (metr-1) as part of the one-carbon cycle. In the mitochondria, vitamin B12 is a cofactor of methylmalonyl coenzyme A mutase. Both genes (metr-1 and mce-1) encoding the metabolic enzymes in which vitamin B12 acts as a cofactor are shown. Red, vitamin B12; Blue, vitamin B6. b CRISPR/Cas9-induced mutations in Ppa-METR-1 and Ppa-MCE-1 with target locations indicated in respective protein domains (sgRNA, arrow). Molecular lesions of isolated mutations via CRISPR/Cas9 are also shown. c, d Requirement of Ppa-metr-1 in transgenerational inheritance of vitamin B12-induced Eu mouth form. c Mean probability of predatory mouth-form in Ppa-metr-1 mutant animals on vitamin B12-supplemented E. coli compared to wild type. d Mean probability of the predatory mouth form in Ppa-metr-1 mutant animals on a Novosphingobium diet and reversal to E. coli compared to the wild type response. e Mean probability of the predatory mouth form in Ppa-mce-1 mutant animals exposed to Novosphingobium and reversal to E. coli compared to wild type response. Final mouth-form frequencies are the mean of at least 10 biological replicates (n = 20 animals per plate). Points represent the mean probability of developing the Eu morph, with error bars reflecting the 95% HDI from the Bayesian model. The yellow area indicates the RSC011 baseline response on E. coli, averaged over 101 generations from a previous study3. See also Supplementary Fig. 2 and Supplementary Table 1. Source data are provided as a Source Data file.
The Eu morph is already induced by 0.1 nM vitamin B12 supplementation
Given that the induction of the Eu morph and the transgenerational memory of the predatory mouth form require different concentrations of vitamin B12, we next wanted to know the minimal amount of vitamin B12 necessary to induce the Eu morph. While our original experiments used a final concentration between 500 and 1500 nM Me-Cbl (Fig. 2), we now reduced the concentration of vitamin B12 for several orders of magnitude (Fig. 4). Supplementation experiments using 250 nM and 125 nM vitamin B12, resulted in an immediate Eu mouth-form induction, but reversal experiments after 5 or 10 generations resulted in an even shorter memory than supplementation with 500 nM vitamin B12. Specifically, the response seen after 250, 125, and 50 nM vitamin B12 lasted for only one generation (Fig. 4a–c and Supplementary Fig. 3a, b). When we reduced the concentration of vitamin B12 even further and supplemented with 10, 5, 2, and 0.1 nM vitamin B12, all four of these concentrations still resulted in a strong induction of the Eu mouth-form (Supplementary Fig. 3c–f). In contrast, worms had lost their memory already in the F5R2 generation (for 50 nM, 0.276 ≤ HDI(θR2) ≤ 0.655, for 10 nM, 0.238 ≤ HDI(θR2) ≤ 0.612, and for 5 nM, 0.27 ≤ HDI(θR2) ≤ 0.645) (Supplementary Fig. 3c–f). Thus, the reduction of vitamin B12 concentration in supplementation experiments results in the loss of TEI of the predatory mouth form, while the induction of the Eu morph remains high. When we reduced the vitamin B12 concentration even further, we found no induction of the predatory mouth form at a concentration of 0.01 nM vitamin B12 (Supplementary Fig. 3g). Instead, worms treated with this amount of vitamin B12 stayed at the baseline level of Eu mouth-form, similar to un-supplemented growth conditions. Therefore, TEI of the predatory mouth form requires a substantially higher vitamin B12 concentration than the initial induction of the Eu mouth form.
Fig. 4: Memory of the induced predatory mouth form after supplementation with different concentrations of vitamin B12, methionine, and folate.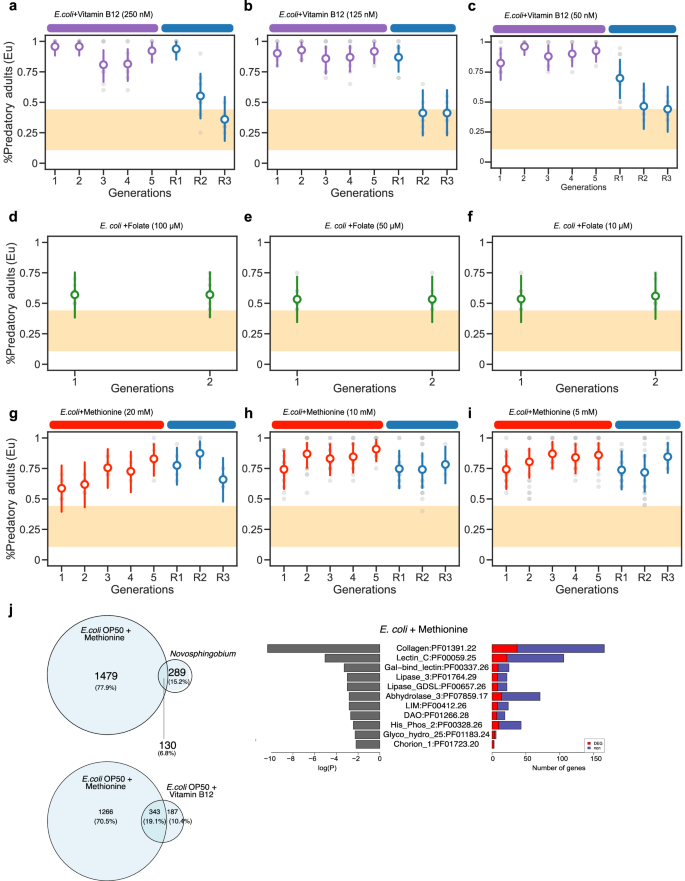
a–c Concentration-dependent effect of vitamin B12 on memory transmission. a Mean probability of predatory mouth-form after a 5-generation exposure to 250 nM vitamin B12 shows Eu memory for two generations. Further reduction of vitamin B12 concentration to b 125 nM, c 50 nM, shows a single generation of Eu memory on un-supplemented E. coli diets. nM, nanomolar. d–f Mean probability of the predatory mouth form on folate-supplemented E. coli plates with d 100 μM, e 50 μM, and f 10 μM working concentration of folate. g–i Mean probability of the predatory mouth form on methionine-supplemented E. coli plates with g 20 mM, h 10 mM, and i 5 mM working concentration of methionine. mM millimolar. μM micromolar. Final mouth-form frequencies are the mean of at least 10 biological replicates (n = 20 animals per plate). Points represent the mean probability of developing the Eu morph, with error bars reflecting the 95% HDI from the Bayesian model. The yellow area indicates the RSC011 baseline response on E. coli, averaged over 101 generations from a previous study3. j Differential gene expression and enrichment analyses from single-worm transcriptomics (SWT). Exposure to methionine-supplemented E. coli showed upregulated overlapping transcripts with Novosphingobium and vitamin B12-supplemented E. coli. Differentially-induced genes are identified relative to worm cultures grown on a standard E. coli diet. The pathways with most significant enrichment (FDR-corrected p value < 0.01) for three generations on methionine-supplemented E. coli are shown. SWT was performed using five independent biological replicates of young adult individuals. See also Supplementary Fig. 3. Source data are provided as a Source Data file.
Memory of the predatory morph requires methionine but not folate
Next, we used supplementation experiments with two major metabolites in the one-carbon cycle (Fig. 3a). First, we used 10, 50, and 100 μM of folate (vitamin B9) for supplementation of standard E. coli plates using similar range of concentrations as a previous study in C. elegans19. However, we found no credible induction of the predatory mouth form (e.g., for 100 μM, 0.384 ≤ HDI(θF2) ≤ 0.755, Fig. 4d–f). Note that this might be due to the micromolar doses of folate that had to be used in these experiments due to solubility. In contrast, methionine supplementation resulted in the induction of the predatory mouth form and subsequent TEI; however, with different dynamics than what is observed after vitamin B12 supplementation. Specifically, supplementation with 5, 10, and 20 mM methionine resulted in the induction of the Eu mouth form after several generations of exposure (Fig. 4g–i). Note that this supplementation does not result in 100% Eu animals similar to supplementation with lower concentrations of vitamin B12 (Fig. 4). However, reversal after five generations of exposure to methionine indicates a TEI of the predatory mouth form that lasts for three generations (Fig. 4g–i). Given that methionine significantly alters osmolarity, we controlled for this effect using sorbitol20. However, sorbitol supplementation did not induce the Eu morph, indicating that the observed response is specific to methionine rather than osmotic changes (Supplementary Fig. 3h). Overall, these results suggest that vitamin B12 might exert its function through methionine in the one-carbon cycle.
Given that vitamin B12 and methionine supplementations can both cause TEI of the predatory morph, we performed single worm transcriptomic analysis of methionine-supplemented worms of day 1 adult individuals. The overlap of upregulated genes between methionine supplementation and Novosphingobium exposure was limited (6.8%), whereas vitamin B12 supplementation showed a larger overlap with Novosphingobium (12.2%), indicating that vitamin B12 recapitulates a larger subset of the bacterial-induced transcriptional program (Fig. 4j). Notably, the overlap between significantly upregulated genes after vitamin B12 and methionine supplementation represents the largest fraction of 19% (Fig. 4j). Together, these patterns suggest that vitamin B12 accounts for a larger fraction of the Novosphingobium-induced transcriptional responses.
Vitamin B12 causes elevated nutrient provisioning
It is important to determine what type of changes act downstream of the original stimulus to cause the inherited effect. This is of particular importance given the different vitamin B12 concentrations necessary to induce the predatory mouth form and its TEI. One possibility would be that vitamin B12 is directly transmitted from the mother to the offspring. Alternatively, TEI might be a consequence of factors acting downstream of vitamin B12. To test for the former possibility, we generated mutants in mrp-5, a gene that was shown in C. elegans to be responsible for the transport of vitamin B12 from mother to the offspring21. Indeed, P. pacificus has a 1:1 ortholog of mrp-5 and we were able to create two mutations in Ppa-mrp-5 that result in frameshifts (Fig. 5a). However, Ppa-mrp-5 mutants show a normal TEI of the predatory mouth form (Fig. 5b, c). These findings suggest that TEI is unlikely to be driven by direct vitamin B12 transmission and instead depends on downstream factors or alternative transport mechanisms.
Fig. 5: Ppa-MRP-5 is dispensable in TEI of the predatory morph and vitellogenin gene cluster organization.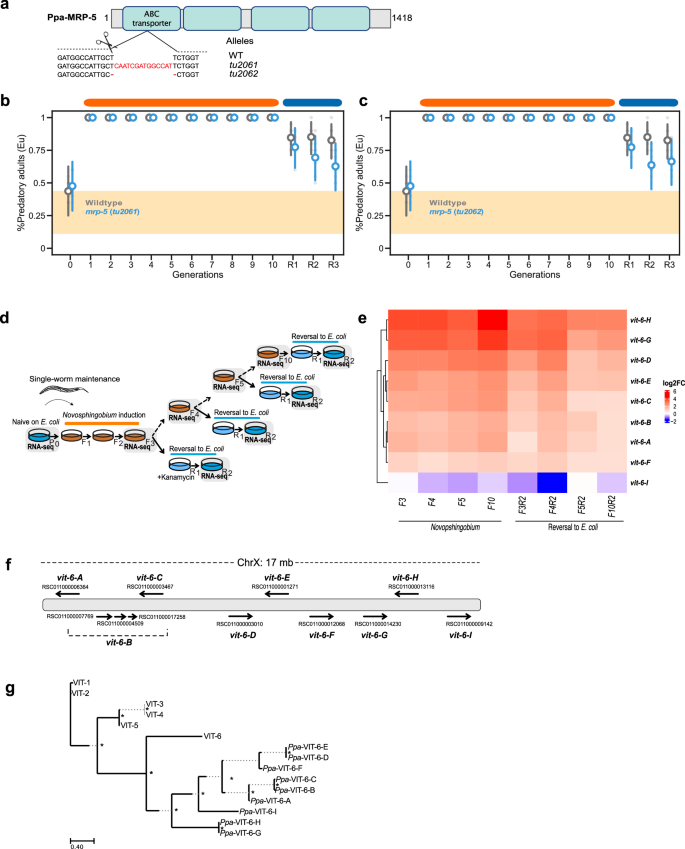
a CRISPR/Cas9-induced mutations in Ppa-MRP-5 with target locations indicated in respective protein domains (sgRNA, arrow). Molecular lesions of isolated mutations via CRISPR/Cas9 are also shown. b, c Mean probability of predatory mouth-form in Ppa-mrp-5 mutant animals (tu2061 and tu2062) on Novosphingobium for 10 generations compared to wild type. Final mouth-form frequencies are the mean of at least 10 biological replicates (n = 20 animals per plate). Points represent the mean probability of developing the Eu morph, with error bars reflecting the 95% HDI from the Bayesian model. The yellow area indicates the RSC011 baseline response on E. coli, averaged over 101 generations from a previous study3. d Graphical representation of the experiment. Grey-highlighted generations indicate populations sent for RNA sequencing. Mixed worm cultures from three agar plates were used to assay independent generational datapoints during Novosphingobium induction and reversal experiments (F3R2, F4R2, F5R2, F10R2). e Vitellogenin expression of all 9 genes under different wildtype conditions: Log2FoldChange (log2FC) values are shown for worms on a Novosphingobium diet (F3, F4, F5, F10) and reversal to E. coli (F3R2, F4R2, F5R2, F10R2). Naïve animals on E. coli (P0) were used as reference to generate log2FC values. f P. pacificus RSC011 has 9 vitellogenin genes all located on the X chromosome, distributed over a 17 Mb range. These genes are named Ppa-vit-6-A to Ppa-vit-6-I based on their location along the chromosome. Note that there are three transcripts that encodes for Ppa-vit-6-B. g Phylogeny of the P. pacificus and C. elegans vitellogenin genes. In C. elegans, six vitellogenin genes show higher sequence divergence with the Cel-vit-6 gene being the most diverse gene copy. Note that it is Cel-vit-6 that is most similar to the individual genes in P. pacificus and also other nematodes. Nodes with bootstrap values of ≥90 are labelled with asterisks (*). Source data are provided as a Source Data file.
In principle, a multigenerational vitamin B12-rich diet could induce a plethora of transcriptional changes in the worm. Therefore, we extended our gene expression analysis to cultures after reversal and compared transcriptional profiles of single naïve, F3, and F5 generation worms on Novosphingobium with F3R2, F4R2, F5R2, and F10R2 generations after reversal to E. coli (Fig. 5d). We used FnR2 rather than FnR1 generations to rule out any effect of the kanamycin treatments in the FnR1 generation. Note that we could not use single worm transcriptomic analyses for this large set of conditions and instead, performed these experiments by harvesting cultures with preferentially adult individuals. We found a consistently strong increase in the expression of vitellogenin genes, encoding the nematode yolk proteins, after Novosphingobium exposure with multiple vitellogenin genes remaining at higher expression levels in comparison to naïve animals (FDR-corrected p value < 0.01) (Fig. 5e). Note that vitellogenin genes were also upregulated in our single worm transcriptomic dataset described above (Fig. 2g), but several of them were not significant. As vitellogenins are transmitted from mothers to offspring, they represent potential candidates for a function in TEI of the predatory mouth form.
P. pacificus shows a more complex vitellogenin composition when compared with C. elegans. While the C. elegans N2 genome contains six vitellogenin genes, P. pacificus RSC011 has 9 genes, all of which are most closely related to Cel-vit-6 (Fig. 5f, g). We therefore named these genes Ppa-vit-6-A to Ppa-vit-6-I based on their location along the chromosome (Fig. 5f, g). While more similar to one another in sequence than the Cel-vit genes, the divergence between the Ppa-vit genes still allows the distinction of individual genes in gene expression analysis (Fig. 5e). We found that 8 of the 9 vitellogenin genes are upregulated upon Novosphingobium exposure, four of which show an increased expression of at least eightfold in the F10 generation (Fig. 5e and Supplementary Data 2). A similar increase in expression was seen in all other tested generations on Novosphingobium. After reversal to E. coli the majority of vitellogenin genes were less expressed than on Novosphingobium; however, expression remained substantially higher than in naïve animals that had never been exposed to Novosphingobium (Fig. 5e). Thus, exposure to a multigenerational Novosphingobium diet and subsequent reversal to E. coli cause elevated vitellogenin transcription resulting in increased nutritional provisioning.
Receptor-mediated endocytosis (rme-2) mutants have no memory
The correlation between elevated vitellogenin expression and the formation of the predatory mouth-form during and after Novosphingobium exposure suggests a role of nutritional provisioning in transgenerational memory. To provide mechanistic evidence for this hypothesis, we investigated the receptor involved in vitellogenin uptake into the germline. In nematodes, vitellogenin is produced in the intestine, and the low-density lipoprotein receptor (LDLR) encoded by rme-2 (receptor-mediated endocytosis protein) was shown to represent the single protein involved in vitellogenin uptake into the germline22,23. In C. elegans, rme-2 mutant embryos contain no detectable yolk and the brood size of homozygous mutant animals is strongly reduced22. In P. pacificus, there is a 1:1 ortholog Ppa-RME-2 with a 55% amino acid sequence similarity (Fig. 6a). Strikingly, we had isolated three mutant alleles of Ppa-rme-2 in our EMS mutagenesis screen for predatory mouth-form Transgenerational-inheritance-defective (Tid) mutants reported previously (Fig. 6a and Supplementary Table 2)3. This finding would be consistent with a role of vitellogenin in memory formation.
Fig. 6: The low-density lipoprotein receptor-related protein RME-2 is required for transgenerational inheritance of the predatory morph.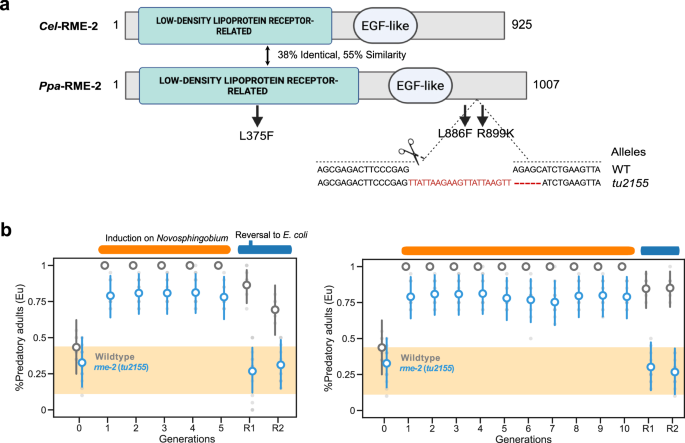
a Domain architecture of C. elegans and P. pacificus RME-2, sequence similarity, and position of molecular lesions in three Ppa-rme-2 mutants are highlighted. Three alleles isolated in a forward genetic EMS mutagenesis screen carry point mutations resulting in amino acid changes as indicated. See also Supplementary Table 2 for the isolated alleles from the genetic screen. We designed a sgRNA in the C-terminal region where two of the original EMS alleles carry their mutation. The frameshift allele tu2215 was used for further analysis. b Mean probability of the predatory mouth form of Ppa-rme-2(tu2215) after a Novosphingobium exposure of 5 (left) and 10 (right) generations. Final mouth-form frequencies are the mean of at least 10 biological replicates (n = 20 animals per plate). Points represent the mean probability of developing the Eu morph, with error bars reflecting the 95% HDI from the Bayesian model. The yellow area indicates the RSC011 baseline response on E. coli, averaged over 101 generations from a previous study3. Source data are provided as a Source Data file.
To rule out the possibility that EMS-induced mutations in genes other than Ppa-rme-2 have resulted in the observed Tid phenotype, we generated an additional knock-out mutation by CRISPR engineering. The Ppa-rme-2(tu2155) allele has a 16 bp insertion that results in a premature stop codon (Fig. 6a). When we tested the Ppa-rme-2(tu2155) allele in a 5-generation Novosphingobium assay, we observed an incomplete induction of the Eu morph for all five generations of Novosphingobium exposures (0.64 ≤ HDI(θF1-tu2155) ≤ 0.928, 0.664 ≤ HDI(θF2) ≤ 0.94\(,\) and 0.628 ≤ HDI(θF5) ≤ 0.921) (Fig. 6b). This result indicates that in the absence of RME-2 and vitellogenin uptake into the germline, the predatory mouth-form can still be induced, which is likely due to other Novosphingobium-derived factors as previously indicated (Fig. 2c). In contrast, after the reversal to E. coli, Ppa-rme-2(tu2155) did not show transgenerational memory of the predatory morph (0.12 ≤ HDI(θF5-tu2155) ≤ 0.431, 0.147 ≤ HDI(θF5R2) ≤ 0.484) (Fig. 6b). Thus, vitellogenin is involved in the formation of transgenerational memory of the predatory mouth-form in P. pacificus indicating a role for nutrient provisioning in this process.




